Map of the Rising Star cave system in Gautong Province South Africa. The yellow dot marks the chamber where Homo naledi fossils were first found; the red one is the site of a new discovery. (Credit: Elliott et al 2021, PaleoAnthropology. Issue 1.64, Fig. 1)
In September 2015 a barrage of publicity announced the remarkable unearthing of the remains of 15 diminutive hominins (see: The ‘star’ hominin of South Africa; September 2015). Subsequently this number rose to at least 24. It was named Homo nadeli, from the floor sediments of an almost inaccessible chamber in the Rising Star cave system of South Africa. In many respects the individuals seemed to share affinities with both australopithecines and the earliest known human, but subsequent dating to 335 to 236 ka proved they were not a ‘missing link’. Many near-complete skeletons suggested that they were similar in many ways to members of the genus Homo. Like H. floresiensis they may have evolved during protracted isolation from their contemporary hominins, perhaps even early modern humans. Nevertheless, it was indeed one of the most dramatic palaeoanthropological discoveries of the 21st century. But its celebrety is as much for the extraordinary efforts of women archaeologists small and lithe enough to wriggle through a maze of extremely narrow cave passages: and the deft PR skills of the leader of the discovering expedition, Lee Berger.
The skeletal remains in the remote chamber were near-complete, with little sign of physical trauma, except gnawing by snails and beetles. Few hominin bones have turned up in the more accessible parts of the cave. One likely explanation was that a living H. naledi group had deliberately carried the bodies through the cave system for burial – at less than 1.5 m tall with a slender build they could have done this far more easily than the modern excavators, but they must have used firebrands to light their way. It was alternatively suggested that a group of them, panicked by large predators, may have scrambled deep into the cave to hide, and suffocated as CO2 built-up to toxic levels, as happens occasionally to trapped speleologists. But another twist has arisen, more than a decade after the discovery.
Applying the large spectrum of modern forensic analysis to such a wealth of bones takes time. One approach, genetic sequencing, demands a great deal of patience. None of the bones preserved ancient DNA, probably because of their age and the high ambient temperatures and humidity of burial at low latitudes. A team led by Palesa Madupe of the Max Planck Institute for Evolutionary Anthropology, Leipzig, Germany then resorted to extracting protein from tooth enamel of 20 individuals (Madupe, P. and 20 others 2026. Proteomic analysis of dental enamel from 20 Homo naledi individuals shows no male markers. Cell, v. 189, p. 1-10; DOI: 10.1016/j.cell.2026.05.044). One dental protein amelogenin (or AMEL) in every analysed hominin species has separate male and female types (AMELX and AMELY), analogous to Y and X chromosomes. Rather than being of mixed sex, the bodies were all women: even the few infants. So, what can that one-in-a-million finding imply?
One of the leaders of the Rising Star team (Lee Berger) had suggested since 2015 that H. naledi had purposefully buried its dead. He now suggests that the species separated sexes in their death rituals. For beings with brains about the size of chimps that seems unlikely to some critics. The earliest evidence for separate mortuary practices only appears in late-Neolithic burials. Moreover, the first evidence for any kind of human inhumation is about 100 ka later than Rising Star at a Neanderthal site in Israel. A possibly more plausible explanation is that H. naledi men were simply too big to reach the burial chamber, i.e. that the species was sexually dimorphic. If so, then why are there no remains of men in more easily accessed parts of the cave system? There remains one other scientific possibility: perhaps H. nadeli had only one type of AMEL and the genes responsible for it – i.e. the L variety was shared between both sexes. Palesa Madupe had previously found AMELY in tooth enamel from a 2 Ma-old South African hominin, so considers that possibility to be highly unlikely. All other approaches seem to be a matter of opinion about cultural practices of which we know little. One seems useful. If H. naledi had a high female to male ratio, perhaps females died at ‘home’, while male hunters were killed while hunting and then eaten by large predators … Further suggestions via Leave a comment.
Hominin fossils referred to as Homo erectus have been found in Africa, Central Europe and China. Those from Africa have also been attributed to H. ergaster and by some to ‘African H. erectus’– a point of lingering dispute and confusion. The African lineage spanned a long time period after appearing around 1.7 Ma ago, possibly to as late as 0.6 Ma. The confusion deepened with the discovery of similar, well-preserved remains at Dmanisi in Georgia that are actually older (1.77–1.85 Ma) than the African specimens. But they are so anatomically diverse that the five skulls might easily be assigned to five different hominin species had they been found at separate locations. Asia is again very odd in an H. erectus context. The species was first proposed in 1891 by Eugene Dubois from remains in sediments oof the Solo River near Trinil in Java – he originally suggested the name Pithecanthropus erectus. Remarkably, the Solo sediments were dated at 53 to 27 ka in 2019, so did Homo erectus co-exist with anatomically modern humans (AMH) on Java? Similarly heavy-browed crania emerged from several sites in China. Curiously, the first complete Denisovan cranium, found near Harbin in China matched H. erectus in the eyebrow department. However, mtDNA from its dental plaque and bone proteomics tie in with those found in fragments from the eponymous Denisova Cave in Siberia and with a fragmentary mandible from Tibet. Without such evidence, and were itnot so young (146 ka), most palaeoanthropologists would probably have called the Harbin individual H. erectus, There are quite a few records of older Chinese hominin crania, dubbed H. erectus on anatomical grounds, including one dated at 1.7 Ma. There are a great many oddities and contradictions that need resolving.
Skulls of Homo erectus from Dmanisi , Georgia (credits; M.S. Ponce de Leon & P.E. Zollkofer, University of Zurich)
The cranium found near Harbin, China belonged to a Denisovan. Credit: Hebei Geo University
On 13 May 2026 a team led by palaeogeneticist Qiaomei Fu of the Chinese Academy of Sciences published data on proteins and amino acids yielded by enamel from six teeth of these ancient Chinese fossils, which emerged from three localities of Middle Pleistocene age (580 to 400 ka) (Fu, Q. and 11 others 2026. Enamel proteins from six Homo erectus specimens across China. Nature, v. 10.1038/s41586-026-10478-8). One is from the Zhoukoudian Cave near Beijing, famous for ‘Peking Man’, the primary reference for the anatomy of Asian H. erectus. Older fossils are unlikely to yield meaningful data of this kind because of chemical degradation; the reason why DNA has so far proved elusive from these specimens. Tooth enamel is extremely durable and can protect proteins and amino acids. Since both are produced by genes on DNA they are proxies for variants of those long-vanished genes
The key protein in the supposed H. erectus teeth isameloblastin which is involved in the formation of tooth enamel. The ameloblastin of all six teeth shared two amino acid variants; one previously unknown in other hominin lineages and perhaps unique to H. erectus, the other has been identified in Denisovans. Fu and colleagues suggest that the original bearers of the teeth – presumed to be H. erectus – had interbred with Denisovans and passed on the second variant gene. In turn that had been passed on to Asian AMH with some of whom Denisovans had interbred; remarkably 21% of living people on the Philippine archipelago carry that gene. The authors go on further to suggest that their findings support the notion that H. erectus was the source by gene-flow for ‘super-archaic’ sections of DNA found in actual Denisovan DNA from one member of that group. That is certainly a possibility, but is not the only one.
Neither the proteomics nor the morphology of the teeth, nor the anatomy of the fossils that accompany them in any way prove that they are from actual 400 ka old Homo erectus individuals. That would require at the very least protein analyses from specimens that definitely pre-date the divergence of Denisovans from Neanderthals about 600 ka ago. Remarkably, proteins have been extracted from a ~1.8 Ma old tooth yielded by the Dmanisi H. erectus site in Georgia, but that failed to reveal anything useful in this context. Maybe future work on older Chinese hominin teeth could resolve the issue. Another hypothesis is that the bearers of the analysed teeth were a population of Denisovans who themselves developed genetic variations rather than inheriting them. Proteomics is at about the same stage in its development as human genomics was before 2010 in the run-up to discovering Neanderthal and Denisovan genomes. But in the case of H. erectus the problem began with biologists’ long record of trying to simplify the natural world, especially fossils, through ‘lumping’ rather than ‘splitting’.
Humans that science has designated as different species were capable of interbreeding over tens and hundreds of thousand years, probably repeatedly and maybe habitually. That fact makes it hard to defend the concept of their speciation. There were few environments where they could not thrive, yet their migrations spread small numbers over vast areas. Continually shifting, isolated populations would diverge genetically and physically, the more so the fewer individuals were banded together. Occasionally populations would meet: an opportunity for celebration, and more, for conscious beings facing the rigours of exploration with neither territory nor resources to defend.
Our hominin ancestors in Africa first fashioned tools about 3.5 Ma ago. Since then regular intake of animal protein through hunting, followed by the later discovery of fire and cooking, may progressively have encouraged the evolution of larger hominin brains. Both behavioural leaps would have reduced the length of the ‘working day’ needed to sustain hominin groups. That would have lengthened opportunities for cognitive reflection. social life and culture, and thus further evolution. They also expanded the opportunities for migration, beginning with Homo ergaster venturing beyond Africa at least 1.8 Ma ago. Hominins evolved to such an extent that several separate species occupied our home world at any one time until about 45 thousand years ago. After that only H. sapiens occupied Africa, Eurasia and Australasia.
Such protracted and meandering evolution and dispersal clearly involved episodic physiological and cultural changes, but all we have to go on are fragmentary fossil remains and artifacts of various kinds. DNA has yet to be extracted from hominin bones older than 400 ka (an early Neanderthal from northern Spain). Though H. sapiens first appeared in Morocco about 300 ka ago, DNA from our species dates back to only 45 ka (western and central Europe). What is today termed ‘ancient’ human DNA, is actually very young and restricted to climate zones where its decomposition has been slow. At present there is little point in analysing fossil material from tropical and subtropical latitudes; the DNA is degraded beyond recovery by even the most up-to-date techniques. Fascinating as discussion of human evolution is, in reality most is merely inferred from comparative anatomy and anthropological interpretation.
By 45 ka the heavy evolutionary lifting had been done, resulting in anatomically modern humans, but we have little, if any, chance of explaining in genetic terms how it was achieved. There has been much speculation about the conditions, particularly climatic ones, which may have driven the changes. During the last 2.6 Ma – the Quaternary Period – global climate has been the most changeable in the last 300 Ma. Ice ages have come and gone, first in 40 ka cycles and during the last million years every 100 ka. Much more rapid changes, such as millennial Dansgaard-Oeschger cycles, appeared during each glacial episode, the last being the Younger Dryas between 12.9 and 11.7 ka. For a long while ideas on the drivers of human evolution have been dominated by those concerning environmental stress. Unsurprisingly, genetic change has also been ascribed to such a Darwinian-ecological cause: adaptability to adversity. To test such a hypothesis requires genetic data, of course. But, except for the climatically more stable Holocene Epoch since 11.7 ka, ancient human genomes are in very short supply.
Renowned researcher into ancient human genetics David Reich of the Harvard Medical School in Boston, USA has collated more than 15 thousand ancient human genomes extracted from the remains of individuals who lived and died in Europe and parts of the Middle East during the last ten thousand years. These have been analysed statistically in the context of ‘directional selection’. This is a type of natural selection that occurs when one version of a gene – an allele – confers an extreme form of a trait. If it proves advantageous it rapidly gets passed on to more descendants than do less advantageous alleles, and thus rises in frequency across a population. This differs from other causes of gene frequency changes, such as human migration, population mixing, and random genetic fluctuations that occur in small populations. Well-known examples of directional selection are rapid changes among European Peppered moths, African cichlid fish, Alaskan Sockeye salmon and Big Cats which change over time in response to variations in their habitats. A human example is a genetic variant that maintains the ability to digest the sugar lactose in milk beyond infancy, which enables many modern Europeans to digest milk throughout their lives. The algorithm needed to separate signs of directional selection from other types of genetic change was developed by Ali Akbari, a computational geneticist also at Harvard Medical School. A recent paper by Akbari, Reich and colleagues in the US, Iran, Germany, and Austria (Akbari, A. and 15 others 2026. Ancient DNA reveals pervasive directional selection across West Eurasia. Nature, advance online publication; DOI: 10.1038/s41586-026-10358-1) seems set dramatically to change the research into recent human evolutionary genetics.
Akbari et al. discovered that directional selection has driven the spread or decline of hundreds of gene variants in human populations throughout Western Europe in the last ten millennia. In particular, selection accelerated with the adoption of farming rather than a hunter-gatherer lifestyle. Among the gene variants are those connected with light skin, red hair, risk of celiac disease – linked to gluten in cereals – susceptibility to gout, resistance to leprosy, baldness, rheumatoid arthritis and alcoholism. There are many more (see Figure 3 in the paper): the team identified 479 gene variants affected by directional selection, some that can be explained by changes in lifestyle, others less explicable and yet more that underlie complex traits such as mental illness and cognition. Some of the variants sprang up and were sustained in the population, others rose and then dwindled. The Neolithic began a period of fundamental life style changes in Europe, summed up as a shift from hunting and foraging to farming of cereals and livestock, as early as about 10 ka ago in what is now Türkiye. The pace of genetic changes of this kind reached a peak around the Bronze Age, perhaps because human activities in Europe became more complex then with the mass migration westwards of Yamnaya horse- and wagon-using people from the steppes to dominate Europe
The shift from small wandering bands to living in settlements was a drastic change from a lifestyle that had continued throughout all previous human history. So, it is hardly surprising that there was a major shift in humans’ genetic makeup. But such a change in human labour was not unique to Europe and is known to have occurred on all inhabited continents, with the exception of Australia, at different times during the Holocene. Other regional genetic databases can be analysed in much the same way, once sufficient ancient DNA is collected in Asia, Africa and the Americas. Yet not much is available. The authors comment: ‘A variant that now correlates to household income or years of schooling [remarkably, there are some!] had to have meant something different in the Stone Age. So these results do not mean that Europeans evolved to be smarter or healthier.’ Moreover, the research results in the paper seem likely to be amplified as the data set is so large and complex.
Followers can now download newly posted annual logs for Human Evolution and Migrations covering the years 2022 to 2025. By downloading them you can get a clear idea of how palaeoanthropology has moved forward since the Covid pandemic.
Enjoy the experience if you have the time and inclination!
For decades, most of the news concerning our deep ancestry emerged from discoveries in sub-Saharan Africa at sites in Zambia, Tanzania, Kenya, South Africa, Ethiopia. The first week of 2026 decisively shifted that focus northwards to Chad and Morocco in two separate publications.
In 2002 ago the world of palaeoanthropology was in turmoil following the first discovery of fragments of what was then thought to be a hominid, or great-ape, cranium in Chad dated at around 7 Ma ago (Brunet, M. and 37 others 2002. A new hominid from the Upper Miocene of Chad, central Africa. Nature, v. 4418, p. 145-151;DOI:10.1038/nature00879). When pieced together the cranium looked like a cross between that of a chimpanzee and an australopithecine. Some suggested that the creature may have been a ‘missing link’ between the hominids and hominins; perhaps the ultimate ancestor of humans. Sahelanthropus tchadensis (nicknamedToumaï or ‘hope of life’ in the local Goran language) was undoubtedly enigmatic. The ‘molecular-clock’ age estimate for the branching of hominins from a common ancestor with chimpanzees was, in 2002, judged to be two million years later the dating of Sahelanthropus, so controversy was inevitable. Another point of contention was the size of Sahelanthropus’s canine teeth: too large for australopithecines and humans, but more appropriate for a gorilla or chimp. Moreover, Toumaï showed no indisputable evidence for having been bipedal. The Chadian site subsequently yielded three lower jaw bones and a collection of teeth, a partial femur (leg bone) and three fragmentary ulnae (forearm bones). The finds suggested that as many as five individuals had been fossilised. The femur gave an unresolved hint of an upright gait, yet the ulnas suggested Toumaï might equally have been arboreal; as could also be said for the australopithecines.
Reconstructed skull of Sahelanthropus tchadensis. (Credit: Didier Descouens, University of Toulouse)
All the limb bones of Toumaïhave now been anatomically compared with those of hominins and apes (Williams S.A. et al. 2026. Earliest evidence of hominin bipedalism in Sahelanthropus tchadensis.Science Advances, v. 12, article eadv0130; DOI: 10.1126/sciadv.adv0130). Scott Williams of New York University and co-workers from other US institutions show that although the leg bones are much the same size as those of chimpanzees, their proportions were more like those of hominins. They also showed features around the knees and hips needed for bipedalism and an insertion point for a tendon for the gluteus maximus muscle (buttock) vital for sustained upright locomotion, similar to the femurs of Orrorin tugenensis (see: Orrorin walked the walk; May 2008) and Ardipithecus ramidus. Unfortunately, an intact Sahelanthropus cranium showing a foramen magnum – where the skull attaches to the spine – continues to elude field workers. Its position distinguishes upright gait definitively.
The second new advance concerns the joint ancestry of Neanderthals, Denisovans and anatomically modern humans (AMH), whose ancient genetics crudely suggest a last common ancestor living between 765 to 550 ka. This had previously been attributed to Homo antecessor found in the Gran Dolina cave at Atapuerca in northern Spain, roughly dated between 950 ka and 770 ka. (Incidentally, Gran Dolina has yielded plausible evidence of cannibalism). A novel possibility stems from hominin fossils excavated from a cave in raised-beach sediments near Casablanca in Morocco (Hublin, JJ. and 28 others, 2026 Early hominins from Morocco basal to the Homo sapiens lineage. Nature, v. 649 ; DOI: 10.1038/s41586-025-09914-y). The fossil-bearing sediments contain evidence for a shift in the Earth’s magnetic field (the Brunhes–Matuyama reversal) dated at 773 ka, much more precisely than the Atapuerca age span for H. antecessor. Jean-Jacques Hublin of CNRS in Paris and his multinational colleagues report that the fossils are similar in age to H. antecessor, yet are morphologically distinct, displaying a combination of primitive traits and of ‘derived features reminiscent of’ later Neanderthal, Denisovan and AMH fossils. The differences and shared features suggest that there may have been genetic exchanges between the Moroccan and Iberian population over a considerable period. The most obvious route would have been across the Straits of Gibraltar, but would have required some kind of water craft. An important question is ‘which population gave rise to the other?’
Artistic reconstruction of a juvenile Homo antecessor, Based on skeletal remains from Gran Dolina Cave
Larger and more robust hominin remains in Algeria dated at 1,000 ka – H. heidelbergensis? – resemble those found near Casablanca. They may have evolved to the latter. Similar possible progenitors to Iberian Homo antecessor have yet to be found in Western Europe. Homo erectus appeared in Georgia and Romania between 2.0 and 1.9 Ma, but the intervening million years or more have yielded no credible European forebears of H. antecessor. For the moment, incursion of a North African population into Europe followed by sustained contact is Hublin et al’s favoured hypothesis, rather than a European origin for Homo antecessor. For Neanderthals and Denisovans to have originated from such an African group, as has been suggested, requires finds of African fossils with plausible resemblance to what are predominantly Eurasian groups. The Iberian population migrated far and wide in Western Europe, as witnessed by stone tools and footprints dating to between 950 to 850 ka in eastern England. So it is equally possible that the Iberian group were progenitors of Neanderthals and Denisovans in Eurasia itself. At least for the moment, ancient genomes of the two H. antecessor groups are unlikely to be found in either Iberian or African fossils of the same antiquity. But, as usual, that will not stifle debate: a resort to the adage ‘absence of evidence is not evidence of absence’ seems appropriate to several research teams!
The oldest anatomically modern human fossils dated at ~300 ka, were also discovered in Morocco (see: Origin of anatomically modern humans, June 2017). Their isolation in the NW corner of the African continent poses a similar conundrum, as since then such beings went on to occupy wide areas of sub-Saharan Africa and then the world.
As early as 3.4 Ma bones with cut marks first appear in Ethiopia suggesting that meat had by then entered the hominin diet. Access to such a rich source of protein has been suggested to have encouraged the evolution of larger hominin brains. By around2.4 Ma ago it may have led to the first known human species (Homo habilis) with a brain larger than those of australopithecines. Homo ergaster, with a significantly larger brain size, first appeared at about 1.8 Ma. As the probable inventor of bifacial stone tools and being the first hominin to leave Africa, H. ergaster needed greater cognitive abilities.It is quite likely that discovery of means to cook food then provided a further boost to human evolution. Cooking unfolds the proteins in meat and also breaks down the constituents of raw vegetables making both more palatable and easier to digest. Also, many potentially nutritious tubers are toxic if not processed and cooked. Another evolutionary advantage is that such an increased uptake of nutrients without needing an increase in successful hunting and foraging reduces the length of the ‘working day’. Fire itself provides warmth, protection from large predators and light, which further increases the time available for social and mental activities.
Being able to cook demands the controlled use of fire. But when was fire first harnessed? After 3 Ma the climate in East Africa cooled to open up dry savannah, prone to wildfires. Finding naturally roasted carcases may have been an incentive for hominins to use smouldering patches to cook meat. The next breakthrough would have been carrying embers to light fires elsewhere. The earliest tentative evidence for such a fire was discovered at Swartkrans in South Africa. Crudely dated between 2 to 1 Ma, it was a reddened patch of soil containing charred, cut marked bones and burnt biface tools. Definite evidence only appears with the burnt teeth of large carp-like fish from a 780 ka site found at Gesher Benot Ya’aqob in Israel. So, archaeological evidence for cooking is very rare. Imagine, then, the excitement of a group of archaeologists from the British Museum and the Natural History Museum in London and several universities in the UK and Netherlands at finding a small, 400 ka-old Neanderthal hearth at Barham in Kent, England containing direct evidence of how the fire was lit (Davis, R. and 14 others 2025.Earliest evidence of making fire. Nature, online advance publication. DOI: 10.1038/s41586-025-09855-6). Apparently, there are signs that fires had been lit at the hearth on twelve or more occasions. So, clearly, the spot was used regularly by Neanderthals.
Striking sparks with flint and pyrite. Credit: Craig Williams, The Trustees of the British Museum
The burnt-earth site yielded fire-cracked flint hand axes and fragments of the mineral pyrite (FeS2). Pyrite, named from the Greek word for ‘fire’ – i.e. ‘fire stone’ – creates showers of sparks when scraped with a hard, sharp tool. Struck into dry grass tinder the sparks cause it to smoulder and then burst into flame when blown on. This approach has been used throughout historic times. Interestingly, pyrite is not found in local rocks and had to have been brought from outcrops of Cretaceous Chalk 15 km away, which is also a major source of flint for stone tools. Not only had the Neanderthal ‘campers’ mastered this fire lighting method, they knew where to get the minerals required. With these skills, they could have lit fires on demand wherever they were; to cook, keep warm, light the night and keep predators at bay. The find is a lucky one, for pyrite eventually oxidises in damp air. The skill may have been acquired long before 400 ka. Yet, as most school children used to know, you can also produce fire, or at least embers and smoke(!), by abrading softwood (a schoolroom desk top) with hardwood (a wooden ruler) … Other abrasive methods are available, but none so handy as a flint-pyrite tinderbox.
See also: Smith, K.N. 2025. Getting lit: This is the oldest evidence of people starting fires. Ars Technica; 10 December 2025
One of the longest-lived hominin species that we know of was Paranthropus boisei, remains of which occur in East African sediments between 2.6 and 1.3 Ma. Others, including our own species, lasted nowhere near as long, except perhaps for Homo erectus who emerged around 1.9 Ma ago and is believed by some to have lingered on in Java until about 112 ka ago. However, when the unresolved muddle in the Middle Pleistocene of similar-looking hominin fossils is eventually unravelled – as now seems to be on the cards – that may limit the range of H. erectus to 1.9 -1.0 Ma. Paranthropoid remains are easily distinguished from those of their contemporary hominins – australopithecines and early species of Homo – being extremely robust compared with the ‘gracile’ members of the human line. They were also bipedal, but their fossil skulls are distinctive: massive teeth and jaws, and a bone crest on top of the cranium to which very powerful chewing muscles were attached. Once regarded as a sort of upright gorilla with vegetarian habits, evidence has accumulated since their first discovery that they may have been far more remarkable.
Reconstruction of a Paranthropus head (Credit: Jerry Humphrey, Pinterest)
The earliest paranthropoid was P. aethiopicus from Ethiopia, dated at around 2.7 to 2.3 Ma, and believed to be the common ancestor of P. boisei and P. robustus found in Tanzania and South Africa respectively. Stone and bone tools associated with paranthropoid remains have been found in South and East Africa, some of which show signs of having been burnt. The connection between paranthropoids and both tool- and fire-making is clearly impossible to verify with certainty, and so too for their known association with australopithecine remains – or even the earliest known humans (Homo habilis) for that matter. Palaeoanthropologists are not likely to find a near-complete skeleton of any of these candidates with a tool grasped in the remains of a hand! The issue can be partly resolved if it can be shown that a fossil hand was capable of making and using tools. The fabled ‘opposable thumb’ that is capable of touching the tips of all four fingers is essential for the necessary ‘precision grip’. Isolated, 2 Ma-old thumb bones probably able to do that were found in the famous Swartkrans Cave in South Africa, but with no clue as to which hominin species had yielded them. In fact had that matter been resolved there and then, it would be not take the hominin story very far, simply because evidence for tool use – tools and cut marks on bone – goes back as far as 3.3 Ma, again with more than one candidate for the usefully endowed hominin species.
The left hand of Paranthropus boisei reconstructed from individual bones, palm-up on the left, palm down on the right. Credit: Mongle et al, Fig 3.
Remarkably, a group of scientists from the US, Canada, Australia, South Africa and Kenya have indeed unearthed from 1.5 Ma sediments on the shore of Lake Turkana in Kenya a near-complete left hand associated with cranial bones and teeth from Paranthropus boisei (Mongle,C.S. and 29 others 2025. New fossils reveal the hand of Paranthropus boisei. Nature v. 647, p. 944–951; DOI: 10.1038/s41586-025-09594-8). It is clear that the P. boisei hand shared some of the manipulative capacity of modern human hands, though it bears some resemblance to gorilla hands. That hand was probably nimble enough to make and use simple stone tools. It would also have had a powerful grip, sufficient for climbing and wielding a large stick. Yet again, it does not indicate which species first adopted tool making and use.
There are several interesting possibilities. It may be that a form of convergent evolution enabled two separate genera to become capable of such skills and the intellect to put them to use: tools, however simple, confer enormous evolutionary advantages. Had the antecedents of humans – presumably a species of Australopithecus – been the first, paranthropoids may have observed and adopted tools or vice versa. Just as possible, the – as yet unknown – common ancestor of both may have made this fundamental leap, which would have benefitted both vegetarian and omnivorous descendants. In that case the physiology of each group may have diverged with their lifestyles. Eating roots and leaves requires considerably more physical effort than getting sufficient protein and fats partly by devouring flesh.
Assigning human fossils older than around 250 ka to different groups of the genus Homo depends entirely on their physical features. That is because ancient DNA has yet to be found and analysed from specimens older than that. The phylogeny of older human remains is also generally restricted to the bones that make up their heads; 21 that are fixed together in the skull and face, plus the moveable lower jaw or mandible. Far more teeth than crania have been discovered and considerable weight is given to differences in human dentition. Teeth are not bones, but they are much more durable, having no fibrous structure and vary a great deal. The main problem for palaeoanthropologists is that living humans are very diverse in their cranial characteristics, and so it is reasonable to infer that all ancient human groups were characterised by such polymorphism, and may have overlapped in their physical appearance. A measure of this is that assigning fossils to anatomically modern humans, i.e. Homo sapiens, relies to a large extent on whether or not their lower mandible juts out to define a chin. All earlier hominins and indeed all other living apes might be regarded as ‘chinless wonders’! This pejorative term suggests dim-wittedness to most people, and anthropologists have had to inure themselves to such crude cultural conjecture.
The extraction, sequencing and comparison of ancient DNA from human fossils since 2010 has revealed that three distinct human species coexisted and interbred in Eurasia. Several well preserved examples of ancient Neanderthals and anatomically modern humans (AMH) have had their DNA sequenced, but a Denisovan genome has only emerged from a few bone fragments from the Denisova Cave in western Siberia. Whereas Neanderthals have well-known robust physical characters, until 2025 palaeoanthropologists had little idea of what Denisovans may have looked like. Then proteins and, most importantly, mitochondrial DNA (mtDNA) were extracted from a very robust skull found around 1931 in Harbin, China, dated at 146 ka. Analysis of the mtDNA and proteins, from dental plaque and bone respectively, reveal that the Harbin skull is likely to be that of a Denisovan. Previously it had been referred to as Homo longi, or ‘Dragon Man’, along with several other very robust Chinese skulls of a variety of ages.
The distorted Yunxian cranium (right) and its reconstruction (middle) [Credit: Guanghui Zhao] compared with the Harbin Denisovan cranium (left) [Hebei Geo University]
The sparse genetic data have been used to suggest the times when the three different coexisting groups diverged. DNA in Y chromosomes from Denisovans and Neanderthals suggest that the two lineages split from a common ancestor around 700 ka ago, whereas Neanderthals and modern humans diverged genetically at about 370 ka. Yet the presence of sections of DNA from both archaic groups in living humans and the discovery that a female Neanderthal from Denisova cave had a Neanderthal mother and a Denisovan father reveals that all three were interfertile when they met and interacted. Such admixture events clearly have implications for earlier humans. There are signs of at least 6 coexisting groups as far back as the Middle Pleistocene (781 to 126 ka), referred to by some as the ‘muddle in the middle’ because such an association has increasingly mystified palaeoanthropologists. A million-year-old, cranium found near Yunxian in Hubei Province, China, distorted by the pressure of sediments in which it was buried, has been digitally reconstructed.
This reconstruction encouraged a team of Chinese scientists, together with Chris Stringer of the UK Museum of Natural History, to undertake a complex statistical study of the Yunxian cranium. Their method compares it with anatomical data for all members of the genus Homo from Eurasia and Africa, i.e. as far back as the 2.4 Ma old H. habilis(Xiabo Feng and 12 others 2025. The phylogenetic position of the Yunxian cranium elucidates the origin of Homo longi and the Denisovans. Science, v. 389, p. 1320-1324; DOI: 10.1126/science.ado9202). The study has produced a plausible framework that suggests that the five large-brained humans known from 800 ka ago – Homo erectus (Asian), H. heidelbergensis, H. longi (Denisovans), H. sapiens, and H. neanderthalensis – began diverging from one another more than a million years ago. The authors regard the Yuxian specimen as an early participant in that evolutionary process. The fact that at least some remained interfertile long after the divergence began suggests that it was part of the earlier human evolutionary process. It is also possible that the repeated morphological divergence may stem from genetic drift. That process involves small populations with limited genetic diversity that are separated from other groups, perhaps by near-extinction in a population bottleneck or as a result of the founder effect when a small group splits from a larger population during migration. The global population of early humans was inevitably very low, and migrations would dilute and fragment each group’s gene pool.
The earliest evidence for migration of humans out of Africa emerged from the discovery of five 1.8 Ma old crania of H. erectus at Dmanisi to the east of the Black Sea in Georgia. similar archaic crania have been found in eastern Eurasia, especially China, at various localities with Early- to Middle Pleistocene dates. The earliest European large-brained humans – 1.2 to 0.8 Ma old H. antecessor from northern Spain – must have migrated a huge distance from either Africa or from eastern Eurasia and may have been a product of the divergence-convergence evolutionary framework suggested by Xiabo Feng and colleagues. Such a framework implies that even earlier members of what became the longi,heidelbergensis, neanderthalensis, and sapiens lineages may await either recognition or discovery elsewhere. But the whole issue raises questions about the widely held view that Homo sapiens first appeared 300 ka ago in North Africa and then populated the rest of that continent. Was that specimen a migrant from Eurasia or from elsewhere in Africa? The model suggested by Xiabo Feng and colleagues is already attracting controversy, but that is nothing new among palaeoanthropologists. Yet it is based on cutting edge phylogeny derived from physical characteristics of hominin fossils: the traditional approach by all palaeobiologists. Such disputes cannot be resolved without ancient DNA or protein assemblages. But neither is a completely hopeless task, for Siberian mammoth teeth have yielded DNA as old as 1.2 Ma and the record is held by genetic material recovered from sediments in Greenland that are up to 2.1 Ma old. The chances of pushing ancient human DNA studies back to the ‘muddle’ in the Middle Pleistocene depend on finding human fossils at high latitudes in sediments of past glacial maxima or very old permafrost, for DNA degrades more rapidly as environmental temperature rises.
Regular readers of Earth-logs will recall that the islands of Indonesia were reached by the archaic humans Homo erectus and H. floresiensis at least a million years ago. Anatomical comparison of their remains suggest that the diminutive H. floresiensis probably evolved from H. erectus under the stress of being stranded on the small, resource-poor island of Flores: a human example of island dwarfism. In fact there are anatomically modern humans (AMH) living on Flores that seem to have evolved dwarfism in the same way since AMH first arrived there between 50 and 5 ka. Incidentally, H. erectus fossils and artefacts were found by Eugene Dubois in the late 19th century at a famous site near Trinil in Java. In 2014, turned out that H. erectus had produced the earliest known art – zig-zag patterns on freshwater clam shells – between 540 and 430 ka ago. The episodic falls in global sea level due to massive accumulations of ice on land during successive Pleistocene glacial episodes aided migration by producing connections between the islands of SE Asia. They created a huge area of low-lying dryland known as ‘Sundaland’. The islands’ colonisation by H. erectus was made easy, perhaps inevitable.
The interconnection of SE Asian islands to form Sundaland (yellow) when sea level was 120 m lower than today. Even at that extreme the island of Sulawesi remained isolated by deep ocean water. Credit: based on Hakim et al Fig 1.
However, Flores and islands further east are separated from those to the west by a narrow but very deep strait. It channels powerful currents that are hazardous to small-boat crossings even today. Most palaeoanthropologists consider the colonisation of Flores by H. erectus most likely to have resulted by accident, reckoning that they were incapable of planning a crossing and building suitable craft. For AMH to have reached New Guinea and Australia around 60 ka ago, they must have developed sturdy craft and sea-faring skills. This paradigm suggests that the evolution of AMH, and thus their eventual occupation of all continents except Antarctica, must have involved a revolutionary ‘leap’ in their cognitive ability just before they left Africa. That view has been popularised by the presenter (Ella Al-Shamahi) of the 2025 BBC Television series Human – now on BBC iPlayer (requires viewers to create a free account) – in its second episode Into the Unknown. [The idea of a cognitive leap that ushered in the almost worldwide migration of anatomically modern humans was launched in 1995 by controversial anthropologist Chris Knight of University College London].
Flaked artefact, about the length of a human thumb, made of chert from excavations at Calio on Sulawesi, dated at 1.02 Ma. Credit: based on Hakim et al Fig 2
The large and peculiarly-shaped island of Sulawesi, also part of Indonesia, is notable for being the location of the earliest known figurative art; a cave painting of a Sulawesi warty pig, dated to at least 45.5 ka ago. Indonesian and Australian archaeologists working at a site near Calio in northern Sulawesi unearthed stone artefacts deep in river-terrace gravels that contain fossils of extinct pigs and dwarf elephants (Hakim, B. and 26 others 2025. Hominins on Sulawesi during the Early Pleistocene. Nature, v. 644;DOI: 10.1038/s41586-025-09348-6). The tools were struck from pebbles of hard fine-grained rocks by flaking to produce sharp edges. A combination of dating techniques – palaeomagnetism, uranium-series and electron-spin resonance – on the terrace sediments and fossils in them yielded ages ranging from 1.04 to 1.48 Ma; far older than the earliest known presence of AMH on the island (73–63 ka). The dates for an early human presence on Sulawesi tally with those from Flores. The tool makers were probably H. erectus. To reach the island from Sundaland at a time when global sea level was 120 m lower than at present would have required crossing more than 50 km of open water. It seems unlikely that such a journey could have been accidental. The migrants would have needed seaworthy craft; possibly rafts. Clearly the AMH crossings to New Guinea around 60 thousand years ago would have been far more daunting. Both land masses would have been below the horizon of any point of departure from the Indonesian archipelago, even with island ‘hopping’. Yet the Sulawesi discovery, combined with the plethora of islands both large and small, suggests that the earlier non-AMH inhabitants of Indonesia potentially could have spread further at times of very low sea level.
The colours of human skin, eyes and hair in living people across the world are determined by variants of genes (alleles) found at the same place on a chromosome. Since chromosomes are inherited from both mother and father, an individual may have the same two alleles (homozygous), or one of each (heterozygous). A dominant allele is always expressed, even if a single copy is present. A recessive allele is only expressed if the individual inherits two copies of it. Most characteristics of individuals result from the interaction of multiple genes, rather than a single gene. A commonly cited example is the coloration of eyes. If we had a single gene for eye colour – that of the iris – that had alleles just for blue (recessive or ‘b’) and one for brown (dominant or ‘B) pigmentation, brown-eyed individuals would have one or two ‘B’ alleles (bB or BB), whereas those with blue eyes would have to have two ‘blue’ alleles (bb). But inheritance is more complicated than that: there are people with green, hazel or grey eyes and even left- and right eyes of different colour. Such examples suggest that there are more than two genes affecting human eye colour, and each must have evolved as a result of mutations. Much the same goes for hair and skin coloration.
A group of scientists from the University of Ferrara in Italy have analysed highly detailed ancient DNA in anatomically modern human remains from Russia (Palaeolithic), Sweden (Mesolithic) and Croatia (Neolithic) to tease out the complexities of pigmentation inheritance. Then they applied a statistical approach learned from that study to predict the likely skin-, eye- and hair pigmentation in 348 less detailed genomes of ancient individuals whose remains date back to 45 Ma ( Silvia Perretti et al, 2025. Inference of human pigmentation from ancient DNA by genotype likelihood. Proceedings of the National Academy of Science, v. 122, article e2502158122; DOI: 10.1073/pnas.2502158122).
An artist’s impression of a Mesolithic woman from southern Denmark (credit: Tom Bjorklund)
All the hunter-gatherer Palaeolithic individuals (12 samples between 45 and 13 ka old) bar one, showed clear signs of dark pigmentation in skin, eyes and hair – the outlier from Russia was probably lighter. Those from the Mesolithic (14 to 4 ka) showed that 11 out of 35 had a light eye colour (Northern Europe, France, and Serbia), but most retained the dark skin and hair expected in descendants of migrants from Africa. Only one 12 ka hunter-gatherer from Sweden had inferred blue eyes, blonde hair, and light skin. The retention of dark pigmentation by European hunter-gatherers who migrated there from Africa has been noted before, using DNA from Mesolithic human remains and in one case from birch resin chewed by a Mesolithic woman. This called into question the hypothesis that high levels of melatonin in skin, which protects indigenous people in Africa from cancers, would result in their producing insufficient vitamin D for good health. That notion supposed that out-of-Africa migrants would quickly evolve paler skin coloration at higher latitudes. It is now known that diets rich in meat, nuts and fungi – staple for hunter-gatherers – provide sufficient vitamin-D for health at high latitudes. A more recent hypothesis is that pale skins may have evolved only after the widespread Neolithic adoption of farming when people came to rely on a diet dominated by cereals that are a poor source of vitamin-D.
However, 132 Neolithic farmers (10 to 4 ka ago) individuals studied by Perretti et al. showed increased diversity in pigmentation, with more frequent light skin tones, yet dark individuals persisted, particularly in southern and eastern Europe. Hair and eye colour showed considerable variability, the earliest sign of red hair showing up in Turkey. Even Copper- and Bronze Age samples ( 113 from 7 to 3 ka) and those from Iron Age Europeans (25 from 3 to 1.7 ka ago) still indicate common retention of dark skin, eyes and hair, although the proportion of lighter pigmentation increased in some regions of Europe. Other analyses of ancient DNA have shown that the Palaeo- and Mesolithic populations of Europe were quickly outnumbered by influx of early farmers, probably from the Anatolian region of modern Turkey, during the Neolithic. The farming lifestyle seems likely to have allowed the numbers of those who practised it to rise beyond the natural environment’s ‘carrying capacity’ for hunter-gatherers. The former inhabitants of Europe may simply have been genetically absorbed within the growing population of farmers. Much the same absorption of earlier groups seems to have happened with the westward migration from the Ukrainian and Russia steppes of the Yamnaya people and culture, culminating in the start of the European Bronze Age that reached western Europe around 2.1 ka, The Yamnaya introduced metal culture, horse-drawn wheeled vehicles and possibly Indo-European language.
So the novel probabilistic approach to ancient DNA by Perretti et al. also casts doubt on the diet-based evolution of light pigmentation at high latitudes. Instead, pulses of large population movements and thus changes in European population genetics probably account for the persistence of abundant evidence for dark pigmentation throughout Europe until historic times. The ‘lightening’ of Europeans’ physiognomy seems to have been vastly more complex than previously believed. Early Europe seems to have been almost bewilderingly diverse, which make a complete mockery of modern chauvinism and racism. The present European genetic ‘melting pot’ is surprisingly similar to that of Europe’s ancient past.
For over a century Chinese scientists have been puzzling over ancient human skulls that show pronounced brow ridges. Some assigned them to Homo, others to species that they believe were unique to China. A widely held view in China was that people now living there evolved directly from them, adhering to the ‘Multiregional Evolution’ hypothesis as opposed to that of ‘Out of Africa’. However, the issue might now have been resolved. In the last few years palaeoanthropologists have begun to suspect that these fossilised crania may have been Denisovans, but none had been subject to genetic and proteomic analysis. The few from Siberia and Tibet that initially proved the existence of Denisovans were very small: just a finger bone and teeth. Out of the blue, teeth in a robust hominin mandible dredged from the Penghu Channel between Taiwan and China yielded protein sequences that matched proteomic data from Denisovan fossils in Denisova Cave and Baishiya Cave in Tibet, suggesting that Denisovans were big and roamed widely in East Asia. In 2021 a near-complete robust cranium came to light that had been found in the 1930s near Harbin in China and hidden – at the time the area was under Japanese military occupation. It emerged only when its finder revealed its location in 2018, shortly before his death. It was provisionally called Homo longi or ‘Dragon Man’. Qiaomei Fu of the Institute of Vertebrate Paleontology and Paleoanthropology in Beijing and her colleagues have made a comprehensive study of the fossil.
The cranium found near Harbin, China belonged to a Denisovan. Credit: Hebei Geo University
It is at least 146 ka old, probably too young to have been H.erectus, but predates the earliest anatomically modern humans to have reached East Asia from Africa (~60 ka ago). The Chinese scientists have developed protein- and DNA extraction techniques akin to those pioneered at the Max Planck Institute for Evolutionary Anthropology in Leipzig. It proved impossible to extract sufficient ancient nuclear DNA from the cranium bone for definitive genomic data to be extracted, but dental plaque (calculus) adhering around the only surviving molar in the upper jaw did yield mitochondrial DNA. The mtDNA matched that found in Siberian Denisovan remains (Qiaomei Fu et al. 2025. Denisovan mitochondrial DNA from dental calculus of the >146,000-year-old Harbin cranium. Cell, v. 188, p. 1–8; DOI: 10.1016/j.cell.2025.05.040). The bone did yield 92 proteins and 122 single amino acid polymorphisms, as well as more than 20 thousand peptides (Qiaomei Fu and 8 others 2025. The proteome of the late Middle Pleistocene Harbin individual. Science, v. 388: DOI: 10.1126/science.adu9677). Again, these established a molecular link with the already known Denisovans, specifically with one of the Denisova Cave specimens. Without the painstaking research of the Chinese team, Denisovans would have been merely a genome and a proteome without much sign of a body! From the massive skull it is clear that they were indeed big people with brains much the same size as those of living people. Estimates based on the Harbin cranium suggest an individual weighing around 100 kg (220 lb or ~15 stone): a real heavyweight or rugby prop!
The work of Qiaomei Fu and her colleagues, plus the earlier, more limited studies by Tsutaya et al., opens a new phase in palaeoanthropology. Denisovans now have a genome and well-preserved parts of an entire head, which may allow the plethora of ancient skulls from China to be anatomically assigned to the species. Moreover, by extracting DNA from dental plaque for the first time they have opened a new route to obtaining genomic material: dental calculus is very much tougher and less porous than bone.
During the Middle Palaeolithic (250 to 30 ka) anatomically modern humans (AMH) and Neanderthals were engaged in new technological developments in Europe and Africa as well as in migration and social interaction. This is reflected in the tools that they left at occupation sites and the fact that most living non-Africans carry Neanderthal DNA. One of the major cultural developments was a novel means of manufacturing stone implements. It developed from the Levallois technique that involved knapping sharp-edged flakes of hard rock from larger blocks or cores. A type of tool first found at a Neanderthal site near La Quina in France is a thick flake of stone with a broad, sharp edge that shows evidence of having been resharpened many times. Most other flake tools seem to have been ‘one-offs’ that were discarded after brief usage. The Quina version was not only durable but seems to have been multipurpose. Analysis of wear patterns on the sharpened edges suggest that they were deployed in carving wood and bone, removing fat and hair from animal hides, and butchery. Such scrapers have been found over a wide area of Europe, the Middle East and NE Asia mostly at Neanderthal sites, including the famous Denisova Cave of southern Siberia that yielded the first Denisovan DNA as well as that of Neanderthals.
Making a typical Quina scraper and related tools. The toolmaker would flake pieces of stone off the core and then carefully shape the Quina scraper. (Image credit: Pei-Yuan Xiao)
Until now, the early humans of East Asia were thought not to have proceeded beyond more rudimentary tools during the Middle Palaeolithic: in fact that archaeological designation hasn’t been applied there. Recent excavations at Longtan Cave in south-west China have forced a complete revision of that view (Ruan, Q.-J., et al. 2025. Quina lithic technology indicates diverse Late Pleistocene human dynamics in East Asia. Proceedings of the National Academy of Sciences, v. 122, article e2418029122; DOI: 10.1073/pnas.2418029122). The Longtan site has yielded more than fifty scrapers and the cores from which they had been struck that clearly suggest the Quina technology had been used there. They occur in cave sediments dated at between 60 and 50 ka. As yet, no human remains have been found in the same level at Longtan, although deeper levels dated at 412 ka have yielded hominin crania, mandibular fragments, and teeth, that have been suggested to be Homo erectus.
Quina type tools in East Asia may previously have been overlooked at other hominin sites in China: re-examination of archived tool collections may show they are in fact widespread. The technology could have been brought in by migrating Neanderthals, or maybe it was invented independently by local East Asian hominins. Because most living people in China carry Denisovan DNA in the genomes so perhaps that group developed the technique before interbreeding with AMH immigrants from the west. Indeed there is no reason to discard the notion that early AMH may have imported the Quina style. A lot of work lies ahead to understand this currently unique culture at Longtan Cave. However, interpretation of another discovery published shortly after that from Longtan has spectacularly ‘stolen the thunder’ of the Qina tools, and it was made in Taiwan …
Right (top) and downward (lower) views of the partial Penghu mandible. Credit: Yousuke Kaifu University of Tokyo, Japan and Chun-Hsiang Chang Tunghai University, Taichung, from Tsutaya et al. Fig. 1 (inset)Taiwan.
About 10 years ago, Taiwanese fishers trawling in the Penghu Channel between Taiwan and China were regularly finding bones in their nets. Between 70 to 10 ka and 190 to 130 ka ago much lower sea level due to continental ice cap formation exposed the Penghu seabed. Animals and humans were thus able to move between the East Asian mainland and what is now Taiwan. The bones brought to the surface included those of elephants, water buffaloes and tigers, but one was clearly a human lower jawbone (mandible). Its shape and large molar teeth are very different from modern human mandibles and molars. A multinational team from Japan, Denmark, Taiwan and Ireland has extracted proteins from the mandible to check its genetic affinities (Tsutaya, T. and 14 others 2025. A male Denisovan mandible from Pleistocene Taiwan. Science, v. 388, p. 176-180; DOI: 10.1126/science.ads3888). Where DNA has not been preserved in bones proteomics is a useful tool, especially if results are matched with other bones that have yielded both DNA and protein sequences. In the case of the Penghu mandible, proteins from its teeth matched those of Denisovans from the Denisova Cave in Siberia which famously yielded the genome of this elusive human group. They also matched proteins from a rib found in Tibet associated with Denisovan mitochondrial DNA in cave sediments that enclosed the bones.
The three sites (Denisova, Baishiya Cave in Tibet and Penghu Channel) that have produced plausible Denisovan specimens span a large range of latitudes and altitudes. This suggests that Denisovans were capable of successful subsistence across much of East Asia. The Penghu mandible and teeth are similar to several hominin specimens from elsewhere in China that hitherto have been attributed to H. erectus. Apart from the Denisovan type locality, most of the sites have yet to be accurately dated. Having been immersed in sea water for thousands of years isotopes used in dating have been contaminated in the Panghu specimen. It can only be guessed to have lived when the seabed from which it was recovered was dry land; i.e. between 70 to 10 ka and 190 to 130 ka. China was undoubtedly occupied by Homo erectus during the early Pleistocene, but much younger fossils have been attributed to that species by Chinese palaeoanthropologists. Could it be that they are in fact Denisovans? Maybe such people independently developed the Quina knapping technique
See also: Marwick, B. 2025. Unknown human species in East Asia used sophisticated tools at the same time Neanderthals did in Europe. Live Science, 31 March 2025; Ashworth. J. 2025. Denisovan jawbone helps to reveal appearance of ancient human species. Natural History Museum News 11 April 2025.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Hominins have been making and using stone tools since at least 3.4 Ma, as shown by cut marks on bones and stone artefacts themselves. I use the sack term ‘hominin’ because the likely makers and users of the oldest tools are either australopithecines or paranthropoids, there being no fossils designates to the genus Homo of late-Pliocene age. So it might seem un-newsworthy to report that the oldest tools deliberately made from bone are now known to occur in 1.5 Ma old sediments from the famous sedimentary sequence at Olduvai Gorge in Tanzania (de la Torre, I and 8 others2025. Systematic bone tool production at 1.5 million years ago. Nature, v. 639; DOI: 10.1038/s41586-025-08652-5). To be clear, there is abundant evidence that hominins had used bones, especially sturdy long bones, for digging perhaps, much earlier in hominin history. Again, paranthropoids have been implicated in their use. The bones found at Olduvai actually show signs of manufacture into useful objects prior to their use: they show clear signs of knapping to produce points and blades. The bones are among the sturdiest known from the Pleistocene, being from elephants and hippos. Before de la Torre and colleagues found what is essentially a bone-tool factory, it was thought that systematic use of bones in such a sophisticated manner only arose between 400 to 250 ka ago among early Homo in Europe. Sadly, fossils of whoever made the tools were not found at the site. Once again, paranthropoids as well as early Homo are known to have cohabited the area at that time.
‘Front, back and side’ views of a 1.5 Ma old tool made from an elephant humerus – its upper foreleg. The scale bar represents 5 cm. (Credit: de la Torre et al.; Fig 3a)
Bifacial Acheulean stone artefacts first appear in the rock record about 300 ka before these bone tools were made. So one idea that the authors put forward is that the same kind of stone knapping technique was transferred to the more abundant massive bones of the East African Pleistocene megafauna (in the absence or rarity of suitable blocks of stone?). But it remains unclear whether or not such tools were simply selected from very large bones smashed to get at their nutritious marrow. The first possibility implies a cultural shift, whereas the latter points simply to expedience. The authors are at pains to point out that the curious million-year gap in the record of bone tools may be ascribed either to the disappearance of bone technology or simply to archaeologists who worked elsewhere having not regarding bone fragments as the products of skills. That applies equally to earlier times, when bones were indeed used, though with not so much in the way of a ‘mental template’. As de la Torre et al. conclude ‘Future research needs to investigate whether similar bone tools were already produced in earlier times, persisted during the Acheulean and eventually evolved into Middle Pleistocene bone bifaces similar in shape, size and technology to their stone counterparts’.
Evidence from Dmanisi in Georgia that Homo erectus may have been the first advanced hominin to leave Africa about 1.8 Ma ago was a big surprise (see: First out of Africa?November 2003). Remains of five individuals included one skull of an aged person who face was so deformed that he or she must have been cared for by others for many years. So, a second surprise from Dmanisi was that human empathy arose far earlier than most people believed. Since 2002 there has been only a single further find of hominin bones of such antiquity, at Longgudong in central China. For the period between 1.0 and 2.0 Ma eight other sites in Eurasia have yielded hominin remains. If finds of stone tools and evidence of deliberate butchery – cut marks on prey animals’ bones – are accepted as tell-tale signs, the Eurasian hominin record is considerably larger, and longer,. There are 11 Eurasian sites that have yielded such evidence – but no hominin remains – that are older than Longgudong: in Russia, China, the Middle East, North Africa and northern India. The oldest, at Masol in northern India is 2.6 Ma old. In January 2025 the earliest European evidence for hominin activity was reported from Grăunceanu in Romania (Curran, S.C. and 15 others 2025. Hominin presence in Eurasia by at least 1.95 million years ago. Nature Communications, v. 16, article 836; DOI: 10.1038/s41467-025-56154-9) in the form of animal bones showing clear signs of butchery, as well as stone tools, but no hominin fossils.
Animal bones showing cut marks from the 1.95 Ma old Grăunceanu site in Romania. (Credit: Curran et al. 2025, Figs 2A and C)
There were stone-tool makers who butchered prey in Africa as early as 3.4 Ma ago (see: Stone tools go even further back; May 2015), but without direct evidence of which hominin was involved. Several possible candidates have been suggested: Australopithecus; Kenyanthropus; Paranthropus. The earliest known African remains of H. erectus have been dated at around 2.0 Ma. So, all that can be said with some certainty about the pre-2 Ma migrants to Eurasia, until fossils of that antiquity are found, is that they were hominins of some kind: maybe advanced australopithecines, paranthropoids or early humans. Those from Longgudong and Dmanisi probably are early Homo erectus, and 2 others (1.7 and 1.6 Ma) from China have been designated similarly. Younger, pre-1.0 Ma Eurasian hominins from Israel, Indonesia, Spain and Turkey are currently un-named at the species level, but are allegedly members of the genus Homo.
So, what can be teased from the early Eurasian hominin finds? Some certainly travelled thousands of kilometres from their assumed origins in Africa, but none penetrated further north than about 50°N. Perhaps they could not cope with winters at higher latitudes, especially during ice ages. To reach as far as eastern and western Eurasia suggests that dispersal following exit from Africa would have taken many generations. There is no reason to suppose continual travel; rather the reverse, staying put in areas with abundant resources while they remained available, and then moving on when they became scarce. Climate cycles, first paced at around 40 ka (early Pleistocene) then at around 100 ka (mid Pleistocene and later), would have been the main drivers for hominin population movements, as it would have been for game and vegetation.
After about 3 Ma the 40 ka climate cyclicity evolved to greater differences in global temperature between glacial and interglacial episodes, and even more so after the mid Pleistocene transition to 100 ka cycles (see Wikipedia entry for the mid-Pleistocene Transition). Thus, it seems likely that chances of survival of dispersed bands of hominins decreased over hundreds of millennia. Could populations have survived in particularly favourable areas; i.e. those at low latitudes? If so did both culture and the hominins themselves evolve? Alternatively, was migration in a series of pulses out of Africa and then dispersal in all directions, most ending in regional extinction? Almost certainly, pressures to leave Africa would have been driven by climate, for instance by increased aridity as global temperatures waned and sea-level falls made travel to Eurasia easier. There may also have been secondary, shorter migrations within Eurasia, again driven by environmental changes. Without more data from newly discovered sites we can go little further. Within the 35 known, pre-1 Ma hominin sites there are two clusters: southern and central China, and the Levant, Turkey and Georgia. Could they yield more developments? A 2016 article in Scientific American about Chinese H. erectus finds makes particularly interesting reading in this regard.
Neanderthals and the elusive Denisovans began to establish permanent Eurasian ranges, after roughly 600 ka ago. Both groups survived until after first contact with waves of anatomically modern humans in the last 100 ka, with whom some interbred before vanishing from the record. However, evidence from the DNA of both groups suggests an interesting possibility. Before the two groups split genetically, their common ancestors (H. heidelbergensis or H. antecessor?) apparently interbred with genetically more ancient Eurasian hominins (see Wikipedia entry for Neanderthal evolution). This intriguing hint suggests that more may be discovered when substantial remains of Denisovans – i.e. more than a few teeth and small bones – are discovered and yield more DNA. My guess is such a future development will stem from analysis of early hominin remains in China, currently regarded as H. erectus. See China discovers landmark human evolution fossils. Xinhua News Agency 9 December 2024)
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
The first anatomically modern humans (AMH) known to have left their remains outside of Africa lived about 200 ka ago in Greece and the Middle East. They were followed by several short-lived migrations that got as far as Europe, leaving very few fossils or artefacts. Over that time Neanderthals were continually present. Migration probably depended on windows of opportunity controlled by pressures from climatic changes in Africa and sea level being low enough to leave their heartland: perhaps as many as 8 or 9 before 70 ka, when continuous migration out of Africa began. The first long-enduring AMH presence in Europe began around 47 ka ago.
For about 7 thousand years thereafter – about 350 generations – AMH and Neanderthals co-occupied Europe. Evidence is growing that the two groups shared technology. After 40 ka there are no tangible signs of Neanderthals other than segments of their DNA that constitute a proportion of the genomes of modern non-African people. They and AMH must have interbred at some time in the last 200 ka until Neanderthals disappeared. In the same week in late 2024 two papers that shed much light on that issue were published in the leading scientific journals, Nature and Science, picked up by the world’s news media. Both stem from research led by researchers at the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany. They focus on new DNA results from the genomes of ancient and living Homo sapiens. One centred on 59 AMH fossils dated between 45 and 2.2 ka and 275 living humans (Iasi, L. M. N. and 6 others 2024. Neanderthal ancestry through time: Insights from genomes of ancient and present-day humans Science, v. 386, p. 1239-1246: DOI: 10.1126/science.adq3010. PDF available by request to leonardo_iasi@eva.mpg.de). The other concerns genomes recovered from seven AMH individuals from the oldest sites in Germany and Czechia. (Sümer, A. P. and 44 others 2024. Earliest modern human genomes constrain timing of Neanderthal admixture. Nature, online article; DOI: 10.1038/s41586-024-08420-x. PDF available by request to arev_suemer@eva.mpg.de ).
Leonardo Iasi and colleagues from the US and UK examined Neanderthal DNA segments found in more than 300 AMH genomes, both ancient and in living people, by many other researchers. Their critical focus was on lengths of such segments. Repeated genetic recombination in the descendants of those individuals who had both AMH and Neanderthal parents results in shortening of the lengths of their inherited Neanderthal DNA segments. That provides insights into the timing and duration of interbreeding. The approach used by Iasi et al. used sophisticated statistics to enrich their analysis of Neanderthal-human gene flow. They were able to show that the vast majority of Neanderthal inheritance stems from a single period of such gene flow into the common ancestors of all living people who originated outside Africa. This genetic interchange seems to have lasted for about 7 thousand years after 50 ka. This tallies quite closely with the period when fossil and cultural evidence supports AMH and Neanderthals having co-occupied Europe.
Reconstruction of the woman whose skull was found at Zlatý kůň, Czechia. Credit: Tom Björklund / Max Planck Institute for Evolutionary Anthropology.
The other study, led by Arev Sümer, has an author list of 44 researchers from Germany, the US, Spain, Australia, Israel, the UK, France, Sweden, Denmark and Czechia. The authors took on a difficult task: extracting full genomes from seven of the oldest AMH fossils found in Europe, six from a cave Ranis in Germany and one from about 230 km away at Zlatý kůň in Czechia. Human bones, dated between 42.2 and 49.5 ka, from the Ranis site had earlier provided mitochondrial DNA that proved them to be AMH. A complete female skull excavated from Czechia site, dated at 45 ka had previously yielded a high quality AMH genome. Interestingly that carried variants associated with dark skin and hair, which perhaps reflect African origins. Neanderthals probably had pale skins and may have passed on to the incomers genes associated with more efficient production of vitamin D in the lower light levels of high latitudes and maybe immunity to some diseases. Both sites contain a distinct range of artefacts known as the Lincombian-Ranisian-Jerzmanowician technocomplex. This culture was once regarded as having been made by Neanderthals, but is now linked by the mtDNA results to early AMH. Such artefacts occur across central and north-western Europe. The bones from both sites are clearly important in addressing the issue of Neanderthal-AMH cultural and familial relationships.
The new, distinct genetic data from the Ranis and Zlatý kůň individuals reveals a mother and her child at Ranis. The female found at Zlatý kůň had a fifth- to sixth-degree genetic relationship with Ranis individuals: she may have been their half first cousin once removed. This suggests a wider range of communications than most people in medieval Europe would have had. The data from both sites suggests that the small Ranis-Zlatý kůň population – estimated at around 200 individuals – diverged late from the main body of AMH who began to populate Asia and Australasia at least 65 ka ago. Their complement of Neanderthal genetic segments seems to have originated during their seven thousand-year presence in Europe. Though they survived through 350 generations it seems that their genetic line was not passed on within and outside of Europe. They died out, perhaps during a sudden cold episode during the climatic decline towards the Last Glacial Maximum. We know that because their particular share of the Neanderthal genome does not crop up in the wider data set used by Iasi et al., neither in Europe and West Asia nor in East Asia. That they survived for so long may well have been due to their genetic inheritance from Neanderthals that made them more resilient to what, for them, was initially an alien environment. It is not over-imaginative to suggest that both populations may have collaborated over this period. But neither survived beyond about 40 ka..
Widely publicised as they have been, the two papers leave much more unanswered than they reveal. Both the AMH-Neanderthal relationship and the general migration out of Africa are shown to be more complex than previously thought by palaeoanthropologists. For a start, the descendants today of migrants who headed east carry more Neanderthal DNA that do living Europeans, and it is different. Where did they interbreed? Possibly in western Asia, but that may never be resolved because warmer conditions tend to degrade genetic material beyond the levels that can be recovered from ancient bones. Also, some living people in the east carry both Neanderthal and Denisovan DNA segments. Research Centres like the Max Planck Institute for Evolutionary Anthropology will clearly offer secure employment for some time yet!
Refugees from the Middle East migrating through Slovenia in 2015. Credit: Britannica
During the Pliocene (5.3 to 2.7 Ma) there evolved a network of various hominins, with their remains scattered across both the northern and southern parts of that continent. The earliest, though somewhat disputed hominin fossil Sahelanthropus tchadensis hails from northern Chad and lived around 7 Ma ago, during the late Miocene, as did a similarly disputed creature from Kenya Orrorin tugenensis (~5.8 Ma). The two were geographically separated by 1500 km, what is now the Sahara desert and the East African Rift System. The suggestion from mtDNA evidence that humans and chimpanzees had a common ancestor, the uncertainty about when it lived (between 13 to 5 Ma) and what it may have looked like, let alone where it lived, makes the notion debateable. There is even a possibility that the common ancestor of humans and the other anthropoid apes may have been European. Its descendants could well have crossed to North Africa when the Mediterranean Sea had been evaporated away to form the thick salt deposits that now lie beneath it: what could be termed the ‘Into Africa’ hypothesis. The better known Pliocene hominins were also widely distributed in the east and south of the African continent. Wandering around was clearly a hominin predilection from their outset. The same can be said about humans in the general sense (genus Homo) during the Early Pleistocene when some of them left Africa for Eurasia. Artifacts dated at 2.1 Ma have been found on the Loess Plateau of western China, and Georgia hosts the earliest human remains known from Eurasia. Since them H. antecessor, heidelbergensis, Neanderthals and Denisovans roamed Eurasia. Then, after about 130 ka, anatomically modern humans progressively populated all continents, except Antarctica, to their geographic extremities and from sea level to 4 km above it.
There is a popular view that curiosity and exploration are endemic and perhaps unique to the human line: ‘It’s in our genes’. But even plants migrate, as do all animal species. So it is best to be wary of a kind of hominin exceptionalism or superior motive force. Before settled agriculture, simply diffusion of populations in search of sustenance could have achieved the enormous migrations undertaken by all hominins: biological resources move and hunter gatherers follow them. The first migration of Homo erectus from Africa to northern China by way of Georgia seems to taken 200 ka at most and covered about ten thousand kilometres: on average a speed of only 50 m per year! That achievement and many others before and later were interwoven with the evolution of brain size, cognitive ability, means of communication and culture. But what were the ultimate drivers? Two recent papers in the journal Nature Communications make empirically-based cases for natural forces driving the movement of people and changes in demography.
The first considers hominin dispersal in the Palaearctic biogeographic realm: the largest of eight originally proposed by Alfred Russel Wallace in the late 19th century that encompasses the whole of Eurasia and North Africa (Zan, J. et al. 2024. Mid-Pleistocene aridity and landscape shifts promoted Palearctic hominin dispersals. Nature Communications, v. 15, article 10279; DOI: 10.1038/s41467-024-54767-0). The Palearctic comprises a wide range of ecosystems: arid to wet, tropical to arctic. After 2 Ma ago, hominins moved to all its parts several times. The approach followed by Zan et al. is to assess the 3.6 Ma record of the thick deposits of dust carried by the perpetual westerly winds that cross Central Asia. This gave rise to the huge (635,000 km2) Loess Plateau. At least 17 separate soil layers in the loess have yielded artefacts during the last 2.1 Ma. The authors radiocarbon dated the successive layers of loess in Tajikistan (286 samples) and the Tarim Basin (244 samples) as precisely as possible, achieving time resolutions of 5 to 10 ka and 10 to 20 ka respectively. To judge variations in climate in these area they also measured the carbon isotopic proportions in organic materials preserved within the layers. Another climate-linked metric that Zan et al. is a time series showing the development of river terraces across Eurasia derived from the earlier work of many geomorphologists. The results from those studies are linked to variations through time in the numbers of archaeological sites across Eurasia that have yielded hominin fossils, stone tools and signs of tool manufacture, many of which have been dated accurately.
The authors use sophisticated statistics to find correlations between times of climatic change and the signs of hominin occupation. Episodes of desertification in Palaearctic Eurasia clearly hindered hominins’ spreading across the continent either from west to east of vice versa. But there were distinct, periodic windows of climatic opportunity for that to happen that coincide with interglacial episodes, whose frequency changed at the Mid Pleistocene Transition (MPT) from about 41 ka to roughly every 100 ka. That was suggested in 2021 to have arisen from an increased roughness of the rock surface over which the great ice sheets of the Northern Hemisphere moved. This suppressed the pace of ice movement so that the 41 ka changes in the tilt of the Earth’s rotational axis could no longer drive climate change during the later Pleistocene, despite the fact that the same astronomical influence continued. The succeeding ~100 ka pulsation may or may not have been paced by the very much weaker influence of Earth changing orbital eccentricity. Whichever, after the MPT climate changes became much more extreme, making human dispersal in the Palearctic realm more problematic. Rather than hominin’s evolution driving them to a ‘Manifest Destiny’ of dominating the world vastly larger and wider inorganic forces corralled and released them so that, eventually, they did.
Much the same conclusion, it seems to me, emerges from a second study that covers the period since ~ 9 ka ago when anatomically modern humans transitioned from a globally dominant hunter-gatherer culture to one of ‘managing’ and dominating ecosystems, physical resources and ultimately the planet itself. (Wirtz, K.W et al. 2024. Multicentennial cycles in continental demography synchronous with solar activity and climate stability. Nature Communications, v. 15, article 10248; DOI: 10.1038/s41467-024-54474-w). Like Zan et al., Kai Wirtz and colleagues from Germany, Ukraine and Ireland base their findings on a vast accumulated number (~180,000) of radiocarbon dates from Holocene archaeological sites from all inhabited continents. The greatest number (>90,000) are from Europe. The authors applied statistical methods to judge human population variations since 11.7 ka in each continental area. Known sites are probably significantly outweighed by signs of human presence that remain hidden, and the diligence of surveys varies from country to country and continent to continent: Britain, the Netherlands and Southern Scandinavia are by far the best surveyed. Given those caveats, clearly this approach gives only a blurred estimate of population dynamics during the Holocene. Nonetheless the data are very interesting.
The changes in population growth rates show distinct cyclicity during the Holocene, which Wirtz et al. suggest are signs of booms and busts in population on all six continents. Matching these records against a large number of climatic time series reveals a correlation. Their chosen metric is variation in solar irradiance: the power per unit area received from the Sun. That has been directly monitored only over a couple of centuries. But ice cores and tree rings contain proxies for solar irradiance in the proportions of the radioactive isotopes 10Be and 14C contained in them respectively. Both are produced by the solar wind of high-energy charged particles (electrons, protons and helium nuclei or alpha particles) penetrating the upper atmosphere. The two isotopes have half-lives long enough for them to remain undecayed and thus detectable for tens of thousand years. Both ice cores and tree rings have decadal to annual time resolutions. Wirtz et al. find that their crude estimates of booms and busts in human populations during the Holocene seem closely to match variations in solar activity measured in this way. Climate stability favours successful subsistence and thus growth in populations. Variable climatic conditions seem to induce subsistence failures and increase mortality, probably through malnutrition.
A nice dialectic clearly emerges from these studies. ‘Boom and bust’ as regards populations in millennial and centennial to decadal terms stem from climate variations. Such cyclical change thus repeatedly hones natural selection among the survivors, both genetically and culturally, increasing their general fitness to their surroundings. Karl Marx and Friedrich Engels would have devoured these data avidly had they emerged in the 19th century. I’m sure they would have suggested from the evidence that something could go badly wrong – negation of negation, if readers care to explore that dialectical law further . . . And indeed that is happening. Humans made ecologically very fit indeed in surviving natural pressures are now stoking up a major climatic hiccup, or rather the culture and institutions that humans have evolved are doing that.
For the last forty thousand years anatomically modern humans have been the only primates living on Planet Earth with a sophisticated culture; i.e. using tools, fire, language, art etcetera. Since Homo sapiens emerged some 300 ka ago, they joined at least two other groups of humans – Neanderthals and Denisovans – and not only shared Eurasia with them, but interbred as well. In fact no hominin group has been truly alone since Pliocene times, which began 5.3 Ma ago. Sometimes up to half a dozen species occupied the habitable areas of Africa. Yet we can never be sure whether or not they bumped into one another. Dates for fossils are generally imprecise; give or take a few thousand years. The evidence is merely that sedimentary strata of roughly the same age in various places have yielded fossils of several hominins, but that co-occupation has never been proved in a single stratum in the same place: until now.
Footprints from Koobi Fora: left – right foot of H. erectus; right – left foot of Paranthropus boisei. Credit: Kevin Hatala. Chatham University
The Koobi Fora area near modern Lake Turkana has been an important, go-to site, courtesy of the Leakey palaeoanthropology dynasty (Louis and Mary, their son and daughter-in-law Richard and Meave, and granddaughter Louise). They discovered five hominin species there dating from 4.2 to 1.4 Ma. So there was a chance that this rich area might prove that two of the species were close neighbours in both space and time. In 2021 Kenyan members of the Turkana Basin Institute based in Nairobi spotted a trackway of human footprints on a bedding surface of sediments that had been deposited about 1.5 Ma ago. Reminiscent of the famous, 2 million years older Laetoli trackway of Australopithecus afarensis in Tanzania, that at Koobi Fora is scientifically just as exciting for it shows footprints of two hominin species Homo erectus and Paranthropus boisei who had walked through wet mud a few centimetres below the surface of Lake Turkana’s ancient predecessor (Hatala, K.G. and 13 others, 2024. Footprint evidence for locomotive diversity and shared habitats among early Pleistocene hominins. Science, v. 386, p. 1004-1010; DOI: 10.1126/science.ado5275). The trackway is littered with the footprints of large birds and contains evidence of zebra.
One set of prints attributed to H. erectus suggest the heels struck the surface first, then the feet rolled forwards before pushing off with the soles: little different from our own, unshod footprints in mud. They are attributed to H. erectus. The others also show a bipedal gait, but different locomotion. The feet that made them were significantly flatter than ours and had a big toe angled away from the smaller toes. They are so different that no close human relative could have made them. The local fossil record includes paranthropoids (Paranthropus boisei), whose fossil foot bones suggest an individual of that speciesmade those prints. It also turns out that a similar, dual walkers’ pattern was found 40 km away in lake sediments of roughly the same age. The two species cohabited the same terrain for a substantial period of time. As regards the Koobi Fora trackway, it seems the two hominins plodded through the mud only a few hours apart at most: they were neighbours.
Artists’ reconstructions of: left – H. erectus; right – Paranthropus boisei. Credits: Yale University, Roman Yevseyev respectively
From their respective anatomies they were very different. Homo erectus was, apart from having massive brow ridges, similar to us. Paranthropus boisei had huge jaws and facial muscles attached to a bony skull crest. So how did they get along? The first was probably omnivorous and actively hunted or scavenged meaty prey: a bifacial axe-wielding hunter-gatherer. Paranthropoids seem to have sought and eaten only vegetable victuals, and some sites preserve bone digging sticks. They were not in competition for foodstuffs and there was no reason for mutual intolerance. Yet they were physically so different that intimate social relations were pretty unlikely. Also their brain sizes were very different, that of P. Boisei’s being far smaller than that of H. erectus , which may not have encouraged intellectual discourse. Both persist in the fossil record for a million years or more. Modern humans, Neanderthals and Denisovans, as we know, sometimes got along swimmingly, possibly because they were cognitively very similar and not so different physically.
Since many hominin fossils are associated with riverine and lake-side environments, it is surprising that more trackways than those of Laetoli and Koobi Fora have been found. Perhaps that is because palaeoanthropologists are generally bent on finding bones and tools! Yet trackways show in a very graphic way how animals behave and interrelate with their environment, for example dinosaurs. Now anthropologists have learned how to spot footprint trace fossils that will change, and enrich the human story.
Aurignacian sculptures: ‘Lion-Man’ and ‘Venus’ from the Hohlenstein-Stadel and Hohle Fels caves in Germany.
The earliest culture (or techno-complex) that can be related to anatomically modern humans (AMH) in Europe is called the Aurignacian. It includes works of art as well as tools made from stone, bone and antler. Perhaps the most famous are the ivory sculptures of ‘Lion-Man’ and Venus of the Hohlenstein-Stadel and Hohle Fels caves in Germany, and also the stunning cave art, of Chauvet Cave in France. Aurignacian artefacts that are dated at 43 to 26 ka occur at sites throughout Europe south of about 52°N. It was this group of people who interacted with the original Neanderthal population of Europe and finally replaced them completely. There is a long standing discussion over who ‘invented’ the stone tools, both human groups apparently having used similar styles of manufacture (Châtelperronian). Likewise, as regards the subsistence methods deployed by each; in one approach Neanderthals may have largely restricted their activities to roughly fixed ranges, whereas the incomers were generally seasonal nomads. As yet it has not been possible to show if the interbreeding between the two, which ancient and modern genetic data show, preceded the Aurignacian influx or continued when the met in Europe. Whatever, Neanderthals as a distinct human group had disappeared from the geological record by 40 ka. (Note that the three thousand years of coexistence is as long as the time between now and the end of the Bronze Age, about 150 generations at least.) But that aspect of European human development is not the only bone of contention about the spread into Europe. How did the Aurignacian people fare during and after their entry into Europe?
Despite continuing discovery of AMH sites in Europe, and reappraisal of long-known ones, there are limits to how much locations, dates, bones and artifacts can tell us. The actual Aurignacian dispersal of people across Europe is confounded by the limited number of proven occupation sites. These were people who, like most hunter gatherers, must have moved continually in response to variations in the supply of resources that depend on changing climatic conditions. They probably travelled ‘light’, occupied many temporary camp sites but few places to which they returned generation after generation. Temporary ‘stopping places’ are difficult to find, showing little more than evidence of fire and a ‘litter’ of shards from retouched stone tools (debitage), together with discarded bones that show marks left by butchery. A group of archaeologists and climate specialists from the University of Cologne, Germany have tried to shed some light on the completely ‘invisible’ aspects of Aurignacian dispersal and subsistence using what they have called – perhaps a tribute to Frank Sinatra! – the ‘Our Way Model’ (Shao, Y. et al. 2024. Reconstruction of human dispersal during Aurignacian on pan-European scale. Nature Communications, v. 15, Article 7406; DOI: 10.1038/s41467-024-51349-y. Click link to download a PDF).
The reality of hunter-gatherer life during a period of repeated rapid change in climate would clearly have been complex and sometimes precarious. To grasp it also needs to take account of human population dynamics as well as climatic and ecological drivers. The team’s basic strategy was to combine climate and archaeological data to model the degree to which human numbers may have fluctuated and the extent and direction of their migration. Three broad factors would have driven both: environmental change; culture – social change, curiosity, technology; and human biology. Really, environmental change is the only one that can be addressed with any degree of precision through records of climate change, such as Greenland ice cores. Archaeological data from known sites should provide some evidence for technological change, but only for two definite phases in Aurignacian culture (43-38 ka and 38-32 ka). Dating of Aurignacian sites establishes some time calibration for episodes of occupation, abandonment and resettlement. Issues of human biology can be addressed to some extent from ancient genetics, where suitable bones are available. However, the ‘Our Way Model’ is driven by climate modelling and archaeology. It outputs an historical estimate of ‘human existence potential’ (HEP) that includes predictions of carbon storage in plants and animals – i.e. potential food resources – expressed as regional population density in Europe. The technical details are complex, but Shao et al.’s conclusions are quite striking.
Maps of estimated anatomically modern human population density during the first six thousand years of Aurignacian migration and palaeoclimate record from the Greenland NGRIP ice core, with shaded warm episodes – red spots indicate the time of the population estimates above. (Credit: Shao et al. Fig. 1)
Climate change in the later stages of cooling towards the last glacial maximum at ~20 ka was cyclical, with ten Dansgaard-Oeschger cold stadial events capable of ‘knocking back’ both population density and the extent of settlement. In the first two millennia expansion from the Levant into the Balkans was slow. From 43 to 41 ka the pace quickened, taking the Aurignacian culture into Western Europe, with an estimate total European AMH population of perhaps 60 thousand. A third phase (41 to 39 ka) shrank the areas and densities of population during a prolonged cold period. The authors suggest that survival was in Alpine refuge areas that AMH people had occupied previously. Starting at around 38 ka, a lengthy climatic warm period allowed the culture to spread to its maximum extent reaching southern Britain and the north and east of the Iberian Peninsula. Perhaps by then the AMH population had evolved better strategies to adapt to increasing frigid conditions. But by that time the Neanderthals had disappeared from Europe freeing up territory and food resources. That too may have contributed to the expansion and the sustenance of an AMH total population of between 80 and 100 thousand during the second phase of the Aurignacian.
It’s as well to remember that this work is based on a model, albeit sophisticated, based on currently known data. Palaeoanthropology is extremely prone to surprises as field- and lab work progresses …
The roof lifted for palaeoanthropologists in October 2004 when news emerged of a fossil from Liang Bua cave on Flores in the Indonesian archipelago. It was an adult female human skull about a third the size of those of anatomically modern humans (see: The little people of Flores, Indonesia; October 2004). Immediately it was dubbed ‘Hobbit’, and from the start controversy raged around this diminutive human. The cave layer contained evidence of fire and sophisticated tools as well as bones of giant rats and minute elephants, presumed to be staple prey for these little people. Despite having brains about the size of a grapefruit – as did australopithecines – the little people challenged our assumptions about intelligence. Preliminary dating from 95 to 17 ka suggested they may have cohabited Indonesia with both H. erectus and AMH. Indeed, modern people of Flores tell legends of the little people they call Ebo Go Go. Like both their ancestors must have crossed treacherous straits between the Indonesian islands, which existed even when global sea level was drawn down by polar icecaps. Once an early suggestion that the original find was the skull of a deformed, microcephalic individual had been refuted by further finds in Flores, scientists turned to natural selection of small stature through living on a small island with limited resources – similar to the tiny elephant Stegodon and other island faunas elsewhere. By 2007, it had become clear from other, similar fossils that they were definitely a distinct species Homo floresiensis (see: Now we can celebrate the ‘Hobbits’! November 2007) with several anatomical similarities to H. erectus. Then more sophisticated dating revealed that the Flores cave sediments containing their fossils and tools spanned 100 to 60 ka, well before AMH reached Indonesia. By 2018 their arrival on Flores, marked by a mandible fragment and 6 teeth in sediments from sediment excavation at Mata Menge 70 km east of Luing Bua, had been pushed back to 773 ka. At the new site stone tools were found in even earlier sediments (1.02 Ma). In 2019 evidence emerged that isolated island evolution in the Philippines had produced similar small descendants (H. luzonensis) by around 67 ka.
Artist’s impression of Homo floresiensis with giant rat. (Credit: Box of Oddities podcast)
The latest development is the finding of a fragment of an adult humerus (an arm bone) in the Mata Menge excavations that had yielded the oldest dates for Homo floresiensis fossils (Kaifu, Y. and 12 others 2024. Early evolution of small body size in Homo floresiensis. Nature Communications, v. 15, article number 6381; DOI: 10.1038/s41467-024-50649-7). Comparing the teeth and arm-bone fragment with an intact adult from Liang Bua suggests that the earliest known ancestors of Homo floresiensis were even smaller. The teeth, albeit much smaller, resemble those of Indonesian specimens of H. erectus. That observation helps to rule out earlier speculation that the tiny people of Flores descended from the earliest humans from Africa (H. habilis) that were about the same size, but more than twice as old (2.3 to 1.7 Ma). The evidence points more plausibly towards their evolution from Asian H. erectus, who arrived in Java around 1.1 million years ago. Having solved the issue of ‘island hopping’ to reach Java a group of Asian H. erectus could have found their way to Flores. That island’s biological resources may not have met the survival requirements of a much larger human ancestor but evolution in isolation kept the arrivals alive. Within 300 ka, and perhaps much less for a small population, survival of smaller offspring allowed them a very long and apparently quite comfortable stay on the island. Though diminished in stature, they demonstrated the survival strategies conferred by being smart.
Charles Darwin famously suggested that humans evolved from apes, and since great apes (chimpanzees, bonobos and gorillas) live in Africa he reckoned it was probably there that the human ‘line’ began. Indeed, the mitochondrial DNA of chimpanzees (Pan troglodytes) is the closest to that of living humans. Palaeoanthropology in Africa has established evolutionary steps during the Pleistocene (2.0 to 0.3 Ma) by early members of the genus Homo: H. habilis, H. ergaster, H. erectus; H. heidelbergensis and the earliest H. sapiens. Members of the last three migrated to Eurasia, beginning around 1.8 Ma with the individuals found at Dmanisi in Georgia. The earliest African hominins emerged through the Late Miocene (7.0 to 5.3 Ma): Sahelanthropus tchadensi, Orrorin tugenensis and Ardipthecus kadabba. Through the Pliocene (5.3 to 2.9 Ma) and earliest Pleistocene two very distinct hominin groups appeared: the ‘gracile’ australopithecines (Ardipithecus ramidus; Australopithecus anamensis; Au. afarensis; Au. africanus; Au. sediba) and the ‘robust’ paranthropoids (Paranthropus aethiopicus; P. robustus and P. boisei). The last of the paranthropoids cohabited East Africa with early homo species until around 1.4 Ma. Most of these species have been covered in Earth-logs and an excellent time line of most hominin and early human fossils is hosted by Wikipedia.
All apes, including ourselves, and fossil examples are members of the Family Hominidae (hominids) which refers to the entire world. A Subfamily (Homininae) refers to African apes, with two Tribes. One, the Gorillini, refers to the two living species of gorilla. The other is the Hominini (hominins) that includes chimpanzees, living humans and all fossils believed to be on the evolutionary line to Homo. The Tribe Hominini is defined to have descended from the common ancestor of modern humans and chimps, and evolved only in Africa. As the definition of hominins stands, it excludes other possibilities! The Miocene of Africa before 7.2 Ma ‘goes cold’ as regards the evolution of hominins. There are, however fossils of other African apes in earlier Miocene strata (8 to 18 Ma) that have been assigned to the Family Hominidae, i.e. hominids, of which more later.
Much has been made of using a ‘molecular clock’ to hint at the length of time since the mtDNA of living humans and chimps began to diverge from their last common ancestor. That is a crude measure at it depends entirely on assuming a fixed rate at which genetic mutation in primates take place. Many factors render it highly uncertain, until ancient DNA is recovered from times before about 400 ka, if ever. The approach suggests a range from 7 to 10 Ma, yet the evolutionary history of chimps based on fossils is practically invisible: the earliest fossil of a member of genus Pan is from the Middle Pleistocene (1.2 to 0.8 Ma) of Kenya. Indeed, we have little if any clue about what such a common ancestor looked like or did. So the course of human evolution relies entirely on the fossil sequence of earlier African hominins and comparing their physical appearances. Each species in the African time line displays two distinctive features. All were bipedal and had small canine teeth. Modern chimps habitually use knuckle walking except when having to cross waterways. As with virtually all other primates, fossil or living, male chimps have large, threatening canines. In the absence of ancient DNA from fossils older than 0.4 Ma these two features present a practical if crude way of assessing to when and where the hominin time line leads.
In 2002 a Polish geologist on holiday at the beach at Trachilos on Crete discovered a trackway on a bedding plane in shallow-marine Miocene sediments. It had been left by what seems to have been a bipedal hominin. Subsequent research was able to date the footprints to about 6.05 Ma. Though younger than Sahelanthropus, the discovery potentially challenges the exclusivity of hominins to Africa. Unsurprisingly, publication of this tentative interpretation drew negative responses from some quarters. But the discovery helped resurrect the notion that Africa may have been colonised in the Miocene by hominins that had evolved in Europe. That had been hinted at by the 1872 excavation of Oreopithecus bamboliifrom an Upper Miocene (~7.6 Ma) lignite mine in Tuscany, Italy – a year after publication of Darwin’s The Descent of Man.
Lignites in Tuscany and Sardinia have since yielded many more specimens, so the species is well documented. Oreopithecus could walk on two legs, its hands were capable of a precision grip and it had relatively small canines. Its Wikipedia entry cautiously refers to it as ‘hominid’ – i.e. lumped with all apes to comply with current taxonomic theory (above). In 2019 another fascinating find was made in a clay pit in Bavaria, Germany. Danuvius guggenmosi lived 11.6 Ma ago and fossilised remains of its leg- and arm bones suggested that it could walk on two legs: it too may have been on the hominin line. But no remains of Danuvius’s skull or teeth have been found. There is now an embarrassment of riches as regards Miocene fossil apes from Europe and the Eastern Mediterranean (Sevim-Erol, A. and 8 others 2023. A new ape from Türkiye and the radiation of late Miocene hominines. Nature Communications Biology, v. 6, article 842.; DOI: 10.1038/s42003-023-05210-5). A number of them closely resemble the earliest fossil hominins of Africa, but most predate the hominin record there by several million years.
Phylogenetic links between fossils assigned to Hominidae found in Africa and north of the Mediterranean Sea. (Credit: Sevim-Erol et al. 2023, Fig 5)
Ayla Sevim-Erol of Ankara University, Turkiye and colleagues from Turkiye, Canada and the Netherlands describe a newly identified Miocene genus, Anadoluvius, which they place in the Subfamily Homininae dated to around 8.7 Ma. Fragments of crania and partial male and female mandibles from Anatolia show that its canines were small and comparable with those of younger African hominins, such as Ardipithecus and Australopithecus. But limb bones are yet to be found. Around the size of a large male chimpanzee, Anadoluvius lived in an ecosystem remarkably like the grasslands and dry forests of modern East Africa, with early species of giraffes, wart hogs, rhinos, diverse antelopes, zebras, elephants, porcupines, hyenas and lion-like carnivores. Sevim-Erol et al. have attempted to trace back hominin evolution further than is possible with African fossils. They compare various skeletal features of different fossils and living genera to assess varying degrees of similarity between each genus, applied to 23 genera. These comprised 7 hominids from the African Miocene, 2 early African hominins (Ardipithecus and Orrorin) and 10 Miocene hominids from Europe and the Eastern Mediterranean. They also assessed similarities with 4 living genera, Homo, orang utan (Pongo), gorilla and chimp (Pan).
The resulting phylogeny shows close morphological links within a cluster (green ‘pools’ on diagram) of non-African hominids with the African hominins, gorillas, humans and chimps. There are less-close relations between that cluster and the earlier Miocene hominids of Africa (blue ‘pool’) and the possible phylogeny of orang utans (orange ‘pool’). Sevim-Erol et al. note that African hominins are clearly more similar and perhaps more closely related to the fossils of Europe and the Eastern Mediterranean than they are to Miocene African hominids. This suggests that evolution among the non-African hominids ceased around the end of the Miocene Epoch north of the Mediterranean Sea. But it may have continued in Africa. Somehow, therefore, it became possible late in Miocene times for hominids to migrate from Europe to Africa. Yet the earlier, phylogenetically isolated African hominids seem to have ‘crashed’ at roughly the same time. Such a complex scenario cannot be supported by phylogenetic studies alone: it needs some kind of ecological impetus.
The Mediterranean Basin at the end of the Miocene Epoch when the only water was in the deepest parts of the basin. (Credit: Wikipedia, Creative Commons)
Following a ‘mild’ tectonic collision between the African continent and the Iberian Peninsula during the late Miocene connection between the Atlantic Ocean and the Mediterranean Sea was blocked from 6.0 to 5.3 Ma. Except for its deepest parts, seawater in the Mediterranean evaporated away to leave thick salt deposits. Rivers, such as the Rhône, Danube, Dneiper and Nile, shed sediments into the exposed basin. For 700 ka the basin was a fertile, sub-sea level plain, connecting Europe and North Africa over and E-W distance of 3860 km. There was little to stop the faunas of Eurasia and Africa migrating and intermingling, at a critical period in the evolution of the Family Hominidae. One genus presented with the opportunity was quite possibly the last common ancestor of all the hominins and chimps. The migratory window vanished at the end of the Miocene when what became the Strait of Gibraltar opened at 5.3 to allow Atlantic water. This resulted in the stupendous Zanclean flood with a flow rate about 1,000 times that of the present-day Amazon River. An animation of these events is worth watching
Until this year, the earliest date recorded for the presence of humans in Europe came from the Sierra de Atapuerca in the Province of Burgos, northern Spain. The Sima del Elefante cave yielded a fossil mandible of a human dubbed Homo antecessor from which an age between 1.2 to 1.1 Ma was estimated from a combination of palaeomagnetism, cosmogenic nuclides and stratigraphy. Stone tools from the Vallonet Cave in southern France are around the same age. There is a time gap of about 200 ka before the next sign of human ventures into Europe, probably coinciding with an extreme ice age. They reappear in the form of stone tools and even footprints that they left between 1.0 to 0.78 Ma in ancient river sediments beneath the crumbling sea cliffs of Happisburgh in Norfolk, England. Although no human fossils were preserved, they too have been assigned to H. antecessor.
Topographic map of Europe (click to see full resolution in a new window). The Carpathian Mountains form an arc surrounding the Pannonian Basin (Hungarian Plains) just below centr. Korolevo and other Homo erectus and H. antecessor sites are marked by red spots (Credit: Wikipedia Commons)
In 1974 Soviet archaeologists discovered a site bearing stone tools by the River Tisza at Korolevo in the Carpathian Mountains close to the borders between Ukraine, Romania and Hungary. Korolevo lies at the northeastern edge of the Pannonian Basin that dominates modern Hungary. Whoever left the tools was on the westward route to a huge, fertile area whose game might support them and their descendants. The route along the Tisza leads to the River Danube and then to its headwaters far to the west. Going eastwards leads to the plains north of the Black Sea and eventually via Georgia to the Levant. On that route lies Dmanisi in Georgia, famous for the site where remains of the first hominins (H. erectus, dated at ~1.8 Ma) to leave Africa were found (see: Consider Homo erectus for what early humans achived). The tools from Korolevo are primitive, but have remained undated since 1974. 50 years on, Roman Garba of the Czech Academy of Sciences with colleagues from Czechia, Ukraine, Germany, Australia, South Africa and Denmark have finally resolved their antiquity (Garba, R. and 12 others 2024. East-to-west human dispersal into Europe 1.4 million years ago. Nature v. 627, p. 805–810; DOI: 10.1038/s41586-024-07151-3). Without fossils it is not possible to decide if the tool makers were H. erectus or H. antecessor.
The method used to date the site is based on radioactive 10Be and 26Al formed from oxygen and silicon in quartz grains by cosmic ray bombardment while the grains are at the surface. Since the half life of 26Al (0.7 Ma) is less than that of 10Be (1.4 Ma), after burial the 26Al/10Be ratio decreases and is a guide to the age of the sediment layer that contains the quartz grains. In this case the ag is quite precise (1.42 ± 0.28 Ma). The decreasing age of H. erectus or H. antecessor sites from the 1.8 Ma of Dmanisi in Georgia in the east, through 1.4 Ma (Korolevo) to 1.2 in Spain and France could mark the slow westward migration of the earliest Europeans. It is tempting to suggest possible routes as Garba et al. have. But such sparse and widely separated sites can yield very little certainty. Indeed, it is equally likely that each known site marks the destination of separate migrations at different times that ended in population collapse. The authors make an interesting point regarding the Korolevo population. They were there at a time when three successive interglacials were significantly warmer than the majority during the Early Pleistocene. Also glacial cycles then had ~41 ka time spans before the transition to 100 ka about 1 Ma ago. Unfortunately, no information about the ecosystem that the migrants exploited is available
The largest volcanic eruption during the 2.5 million year evolution of the genius Homo, about 74 thousand years (ka) ago, formed a huge caldera in Sumatra, now filled by Lake Toba. A series of explosions lasting just 9 to 14 days was forceful enough to blast between 2,800 to 6,000 km3 of rocky debris from the crust. An estimated 800 km3 was in the form of fine volcanic ash that blanketed South Asia to a depth of 15 cm. Thin ash layers containing shards of glass from Toba occur in marine sediments beneath the Indian Ocean, the Arabian and South China Seas. Some occur as far off as sediments on the floor of Lake Malawi in southern Africa. A ‘spike’ of sulfates is present at around 74 ka in a Greenland ice core too. Stratospheric fine dust and sulfate aerosols from Toba probably caused global cooling of up to 3.5 °C over a modelled 5 years following the eruption. To make matters worse, this severe ‘volcanic winter’ occurred during a climatic transition from warm to cold caused by changes in ocean circulation and falling atmospheric CO2 concentration, known as a Dansgaard-Oeschger event.
There had been short-lived migrations of modern humans out of Africa into the Levant since about 185 ka. However, studies of the mitochondrial DNA (mtDNA) of living humans in Eurasia and Australasia suggest that permanent migration began about 60 ka ago. Another outcome of the mtDNA analysis is that the genetic diversity of living humans is surprisingly low. This suggests that human genetic diversity may have been sharply reduced globally roughly around the time of the Toba eruption. This implies a population bottleneck with the number of humans alive at the time to the order of a few tens of thousands (see also: Toba ash and calibrating the Pleistocene record; December 2012). Could such a major genetic ‘pruning’ have happened in Africa? Over six field seasons, a large team of geoscientists and archaeologists drawn from the USA, Ethiopia, China, France and South Africa have excavated a rich Palaeolithic site in the valley of the Shinfa River, a tributary of the Blue Nile in western Ethiopia. Microscopic studies of the sediments enclosing the site yielded glass shards whose chemistry closely matches those in Toba ash, thereby providing an extremely precise date for the human occupation of the site: during the Toba eruption itself (Kappelman, Y. and 63 others 2024. Adaptive foraging behaviours in the Horn of Africa during Toba supereruption. Nature, v. 627; DOI: 10.1038/s41586-024-07208-3).
Selection of possible arrowheads from the Shinfa River site (Credit: Kappelman et al.; Blue Nile Survey Project)
The artifacts and bones of what these modern humans ate suggest a remarkable scenario for how they lived. Stone tools are finely worked from local basalt lava, quartz and flint-like chalcedony found in cavities in lava flows. Many of them are small, sharp triangular points, some of which show features consistent with their use as projectile tips that fractured on impact; they may be arrowheads, indeed the earliest known. Bones found at the site are key pointers to their diet. They are from a wide variety of animal, roughly similar to those living in the area at present: from monkeys to giraffe, guinea fowl to ostrich, and even frogs. There are remains of many fish and freshwater molluscs. Although there are no traces of plant foods, clearly those people who loved through the distant effects of Toba were well fed. Although a period of global cooling may have increased aridity at tropical latitudes in Africa, the campers were able to devise efficient strategies to obtain victuals. During wet seasons they lived off terrestrial prey animals, and during the driest times ate fish from pools in the river valley. These are hardly conditions likely to devastate their numbers, and the people seem to have been technologically flexible. Similar observations were made at the Pinnacle Point site in far-off South Africa in 2018, where Toba ash is also present. Both sites refute any retardation of human cultural progress 74 ka ago. Rather the opposite: people may have been spurred to innovation, and the new strategies may have allowed them to migrate more efficiently, perhaps along seasonal drainages. In this case that would have led them or their descendants to the Nile and a direct route to Eurasia; along ‘blue highway’ corridors as Kappelman et al. suggest.
Yet the population bottleneck implied by mtDNA analyses is only vaguely dated: it may have been well before or well after Toba. Moreover, there is a 10 ka gap between Toba and the earliest accurately dated migrants who left Africa – the first Australians at about 65 ka. However, note that there is inconclusive evidence that modern humans may have occupied Sumatra by the time of the eruption. Much closer to the site of the eruption in southeast India, stone artifacts have been found below and above the 74 ka datum marked by the thick Toba Ash. Whether these were discarded by anatomically modern humans or earlier migrants such as Homo erectus remains unresolved. Either way, at that site there is no evidence for any mass die-off, even though conditions must have been pretty dreadful while the ash fell. But that probably only lasted for little more than a month. If the migrants did suffer very high losses to decrease the genetic diversity of the survivors, it seems just as likely to have been due to attrition on an extremely lengthy trek, with little likelihood of tangible evidence surviving. Alternatively, the out-of-Africa migrants may have been small in number and not fully representative of the genetic richness of the Africans who stayed put: a few tens of thousand migrants may not have been very diverse from the outset.
February 2024 was a notable month for discoveries about ancient technology: first that of an ancient tool probably used in rope making and now signs of the use of a composite ‘plastic’ material in stone-tool hafts. Both are from Neanderthal sites in France, the first dated around 52 to 41 ka and the second in the Le Moustier rock shelters of the Dordogne – the type locality for the Mousterian culture associated with Neanderthals (Schmidt, P. et al. 2024. Ochre-based compound adhesives at the Mousterian type-site document complex cognition and high investment. Science Advances, v. 10, article ead10822; DOI: 10.1126/sciadv.adl0822), dated at around 56 to 40 ka. The second discovery resulted from the first detailed analysis of unstudied artifacts unearthed from Le Moustier in 1907 by Swiss archaeologist Otto Hauser that had been tucked away in a Berlin Museum.
Patrick Schmidt of the University of Tubingen in Germany and colleagues from Germany, the US and Kazakhstan identified stone artifacts that show traces of red and yellow colorants. At first sight it could be suggested that they are decorations of some kind. However, they coat only parts of the stone flakes and are sharply distinct from the fresh rock surface and the sharpest edges. Another feature discovered during chemical analysis is that the colour is due to iron hydroxides (goethite) but this ochre is mixed with natural bitumen: the coating is a composite of an adhesive and filler not far different from what can be purchased in any hardware store.
LEFT: Stone flake from the Le Moustier site in France, partly coated with a reddish iron-rich colorant. RIGHT: Experimental stone flakes with 55:44 mix of goethite and bitumen (top) and pure bitumen (bottom) being handled. (Credit: Schmidt et al. Figs 1A, 3).
The authors tested the properties of the mixtures against those of bitumen alone – an adhesive known to have been used along with various tree resins to haft blades to spears in earlier times. In particular they examined the results of ‘cooking’ the substances. Whether unheated or ‘cooked’ a mixture of ochre and bitumen is up to three times stronger than pure bitumen. A further advantage is that the mixed ingredients are not sticky when cooked and cooled, whereas bitumen remains sticky, as the illustration clearly shows. Anyone who has handled a stone blade realises how sharp they are, and not just around the cutting edges. So Schmidt and colleagues tried to use the composite material as a protective handle when stone flake tools were gripped for cutting or carving. The composite handles worked well on scrapers and blades, even in the softer, ‘uncooked’ form
Similar composite adhesives are known from older sites in Africa associated with anatomically modern humans, but not for this particular, very practical use. It is perhaps possible that the use of bitumen mixed with ochre was brought into Europe by AMH migrants and adopted by Neanderthals who came into contact with them. Yet the limestones of the Dordogne valley yield both bitumen in liquid and solid forms, and ochers are easily found because of their striking colours. Long exposure of petroleum seeps drives off lighter petroleum compounds to leave solid residues that can be melted easily to tarry consistency. So there is every reason to believe that Neanderthals developed this technology unaided. As Schmidt has commented, “Compound adhesives are considered to be among the first expressions of the modern cognitive processes that are still active today”.
Even at my age, if I rummage through pockets of various bits of outdoor clothing there’s a good chance I’ll find a handy length of string that I have scavenged at some time. It’s a just-in-case thing, which I learned from my father and grandad. One can hardly imagine a hunter-gatherer not having string or lengths of sinew for that very reason. Cordage has many other uses than merely securing something: bags, mats, nets, snares, fabric, baskets, huts made of sticks and fronds, and even watercraft. Yet archaeological evidence for twine is exceedingly rare. The oldest known string – made of bark fibres – was found wrapped around a stone tool at a 52 to 41 ka Neanderthal site in the Rhône valley 120 km north-west of Marseille. Rope is somewhat more difficult to make as it requires twisting together several lengths of simpler cordage. Once that skill is cracked a rope maker is on the verge of engineering!
The reassembled rope-making tool from Hohle Fels Cave (Credit: Conard & Rots, Fig 2)
In 2015 archaeologists unearthed several pieces of worked mammoth ivory from the Hohle Fels Cave in SW Germany. They were dated to between 40 to 35 ka and associated with Aurignacian stone tools made by modern humans. Fifteen pieces could be fitted together to yield a 20 cm long ‘baton’. First believed to be some kind of ritual object, the fact that 4 circular holes had been bored through the ‘baton’ suggested it must have had some practical use, perhaps for straightening wooden shafts. Then it became clear that each hole was surrounded by spirals of carefully carved, V-shaped notches. Nicholas Conard and Veerle Rots of the University of Tubingen realised that the object may have been used for making rope using a technique known from the Egyptian pharaonic period into medieval times (Conard, N.J. & Rots, V. 2024. Rope making in the Aurignacian of Central Europe more than 35,000 years ago. Science Advances, v. 10, article adh5217; DOI: 10.1126/sciadv.adh5217).
Frame from a movie showing how the tool may have been used to make ropes. The three ‘feeders’ twist foliage clockwise whereas the fourth pulls and imparts an anticlockwise twist to the three stands. (Credit: Conard & Rots, Supplementary material, Fig S15)
After a little practice, four people were able to make sturdy rope using a replica of the tool. Three twisted together fibrous materials, such as the stems and leaves of bulrushes (Typha), and pushed the rough cordage through the intact holes. A fourth person pulled the cordage through and counter-twisted the three strands into rope about 1.5 cm thick – thicker rope would also have required a tool with more holes and more operators. The spiral grooves maintained the initial clockwise rotation of each strand of cordage, so that when all three were twisted together in an anticlockwise sense the counter rotation held the rope together firmly. Remarkably, the small team were able to produce 5 m of rope in 10 minutes. Other common kinds of fibrous plant material, such as linden and willow were used successfully. Incidentally, the tool squeezed edible starch from the foliage of bulrushes. But it seems that this particular rope-making took only performed well for coarse materials. Making rope from finer firbres, such as animal sinew, nettle, flax and hemp would probably have required another design with smaller holes.
Good illustrations of self publicity and soaring ambition are the private space programmes of oligarchs Elon Musk (SpaceX), Jeff Bezos (Blue Origin) and Richard Branson (Virgin Galactic). For a cool US$65 million a ‘civilian’ can get a trip to the International Space Station on SpaceX; a one-hour suborbital flight on Blue Origin will cost US$300,000, with luck having Bezos as a companion; a reservation on Virgin Galactic for a 1 hour trip to the ‘edge of space’ (~100 km up) now costs US$624,000. It’s a tourist trip for the very, very rich only … but even the long-dead can go … or bits of them. On 8 September 2023 aboard Virgin Galactic flight Tim Nash, a South African billionaire had in his pocket a sturdy tube containing a thumb bone of Homo nalediand the collarbone of Australopithecus sediba. Nash reportedly said afterwards, “I am humbled and honoured to represent South Africa and all of humankind as I carry these precious representations of our collective ancestors”.
Reconstructed head of a somewhat annoyed Homo naledi. Credit: John Gurche, Mark Thiessen, National Geographic.
Nash was entrusted with these unique fossils by Lee Berger, Professor in Palaeoanthropology at Witwatersrand University, South Africa and a National Geographic Explorer-in-Residence. Berger recovered fossils of both species from limestone caves in the UNESCO World Heritage Site grandly named the Cradle of Humankind near Johannesburg. He is no stranger to controversy, and this venture cooked up with Nash seems to aim at promotion of South African achievements rather than having any scientific purpose. It has backfired spectacularly (see: McKie, R. 2023. ‘Callous, reckless, unethical’: scientists in row over rare fossils flown into space. The Observer, 22 October 2023). Comments from the anthropological world, six national and international bodies and perhaps the leading hominin specialist Professor Chris Stringer of the Natural History Museum in London include the words and phrases “callous”, “unethical”, “extraordinarily poorly thought-out”, “a publicity stunt”, “reckless” and “utterly irresponsible”. The caper breaks the South African, indeed international, scientific rule that fossils can only be allowed to travel for scientific purposes, applied consistently by similarly hominin-rich African countries such as Ethiopia, Kenya and Tanzania.
But, Hey, that’s how you get on in the world … isn’t it?



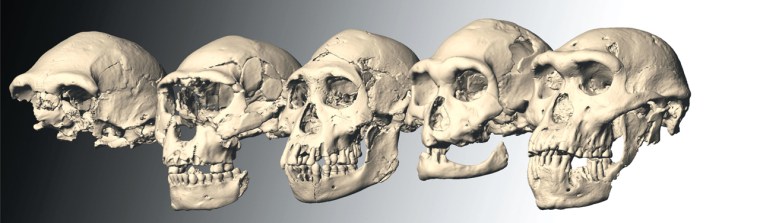










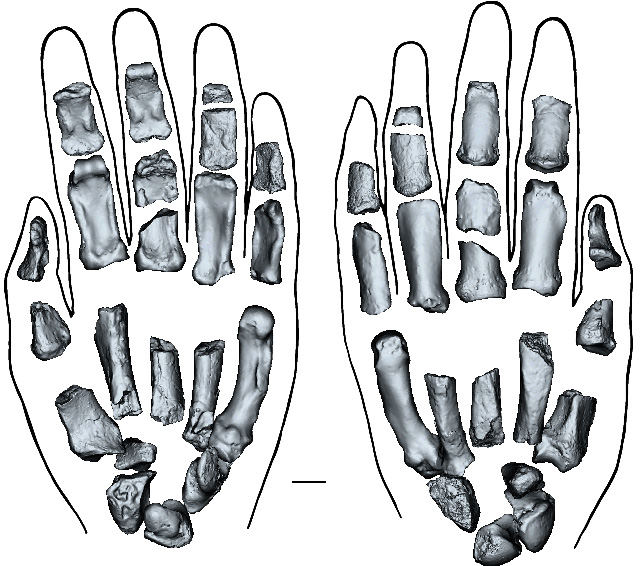



























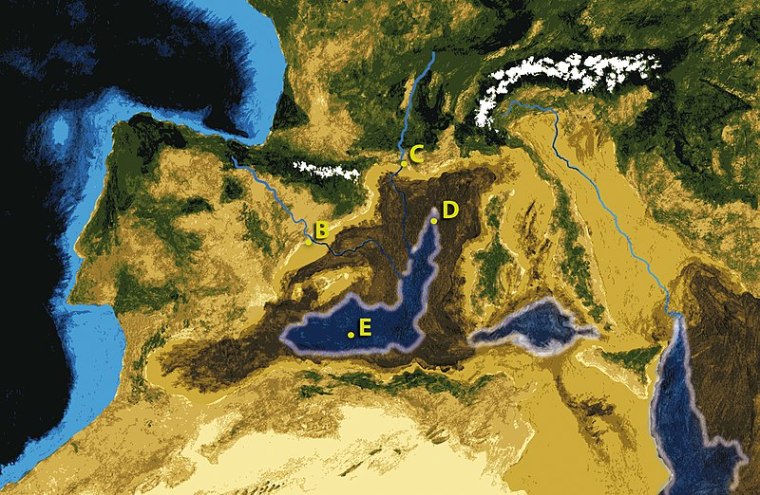













{kind=link}