During the past 539 Ma (the Phanerozoic Eon) Earth’s geological history saw the explosion of rapidly evolving life in the oceans and on the land. The pace of that evolution swung up and down through a complex sequence of extinctions and adaptive radiations. They resulted from many intertwined inorganic changes: tectonics; impacts; igneous events; global climate change; atmosphere and sea-water composition. Although palaeoclimatic knowledge has become ever more detailed over the last few decades, its most important record, the varying temperature of the land surface and oceans, is lacking in precision. The timing of climatic events is not the issue, but the magnitude of changes in global mean surface temperature. The latter is largely down to the main tool in assessing past temperatures: the isotopic composition of oxygen (δ18O) in marine fossils. In particular, the record for the Lower Palaeozoic has remained stubbornly odd. In the Cambrian and Ordovician Periods it implies that low-latitude seawater temperatures reached levels of 40 to 50 °C, that seem literally life threatening: phytoplankton at the base of modern marine ecosystems die at water temperatures above 35°C. Yet the fossil record is teeming throughout the Lower Palaeozoic at all latitudes. Some manner of imprecision in the oxygen-isotope method gives the impression of wild fluctuations and a dramatic overall cooling of the planet through the Phanerozoic: the temperature record as it stands seems implausible.
The carbonate-silicate cycle within the longer-term carbon cycle. Source: Wikimedia Commons
A group of palaeoclimatologists from China, the UK, Australia and the US have combined a variety of geochemical proxies, sedimentary records and climate modelling to correct the marine-carbonate δ18O record (Zheng, D. and 12 others 2026. Tight regulation of Earth’s long-term temperature over Phanerozoic time. Nature Communications, in press 4 May 2026; DOI: 10.1038/s41467-026-72672-6). Their approach is based on a chemical index of alteration (CIA), i.e. a measure of the degree of chemical weathering of the source for sedimentary rocks. The CIA compares their content of immobile aluminium oxide (Al2O3) with calcium, sodium and potassium oxides that are more easily moved in solution. Analyses of recent river sediments show a positive correlation between CIA and local temperature, so CIA in ancient sedimentary rocks is a potential proxy for the ambient temperature of the region from which those sediments were derived. The CIA also depends on other factors, such as the intensity of physical erosion and transport. However, allowing for these factors in modern environments does not affect the correlation with ambient temperature: the method remains robust. The geochemical data from sedimentary rocks required to use CIA as an independent check on O-isotope derived temperature are available in abundance from all continents for most of the Phanerozoic.
The study by Zheng et al. suggests that throughout the Phanerozoic global mean temperature remained consistently within the 10 to 30°C range. Thus Palaeozoic ocean temperatures were comparable with those of the succeeding Mesozoic and Cenozoic Eras. The team concludes that various negative feedback processes inherent in the Earth System have been able to regulate its surface temperature through the Phanerozoic. The most important of these is climate-dependent silicate weathering in which acidic rain – produced by CO2 dissolved from the atmosphere – breaks down silicates to yield dissolved bicarbonate ions that combine with calcium and magnesium ions to precipitate carbonates. Such a process draws down the main greenhouse gas from the atmosphere. There are other aspects of the carbon cycle that also draw down atmospheric CO2 and reduce the greenhouse effect, such as burial of organic debris. Tectonics also shapes climate by modulating both silicate weathering and CO2 emissions from volcanic activity.
It should be emphasised that anthropogenic global warming is proceeding at a far higher rate than natural negative feedback processes. We simply cannot rely on silicate weathering to reverse whatever climatic outcome results from what the current global economy does so very quickly. Yet the findings by Zheng et al. do seem likely to force a change in thinking about climate change on a geological timescale.
Our hominin ancestors in Africa first fashioned tools about 3.5 Ma ago. Since then regular intake of animal protein through hunting, followed by the later discovery of fire and cooking, may progressively have encouraged the evolution of larger hominin brains. Both behavioural leaps would have reduced the length of the ‘working day’ needed to sustain hominin groups. That would have lengthened opportunities for cognitive reflection. social life and culture, and thus further evolution. They also expanded the opportunities for migration, beginning with Homo ergaster venturing beyond Africa at least 1.8 Ma ago. Hominins evolved to such an extent that several separate species occupied our home world at any one time until about 45 thousand years ago. After that only H. sapiens occupied Africa, Eurasia and Australasia.
Such protracted and meandering evolution and dispersal clearly involved episodic physiological and cultural changes, but all we have to go on are fragmentary fossil remains and artifacts of various kinds. DNA has yet to be extracted from hominin bones older than 400 ka (an early Neanderthal from northern Spain). Though H. sapiens first appeared in Morocco about 300 ka ago, DNA from our species dates back to only 45 ka (western and central Europe). What is today termed ‘ancient’ human DNA, is actually very young and restricted to climate zones where its decomposition has been slow. At present there is little point in analysing fossil material from tropical and subtropical latitudes; the DNA is degraded beyond recovery by even the most up-to-date techniques. Fascinating as discussion of human evolution is, in reality most is merely inferred from comparative anatomy and anthropological interpretation.
By 45 ka the heavy evolutionary lifting had been done, resulting in anatomically modern humans, but we have little, if any, chance of explaining in genetic terms how it was achieved. There has been much speculation about the conditions, particularly climatic ones, which may have driven the changes. During the last 2.6 Ma – the Quaternary Period – global climate has been the most changeable in the last 300 Ma. Ice ages have come and gone, first in 40 ka cycles and during the last million years every 100 ka. Much more rapid changes, such as millennial Dansgaard-Oeschger cycles, appeared during each glacial episode, the last being the Younger Dryas between 12.9 and 11.7 ka. For a long while ideas on the drivers of human evolution have been dominated by those concerning environmental stress. Unsurprisingly, genetic change has also been ascribed to such a Darwinian-ecological cause: adaptability to adversity. To test such a hypothesis requires genetic data, of course. But, except for the climatically more stable Holocene Epoch since 11.7 ka, ancient human genomes are in very short supply.
Renowned researcher into ancient human genetics David Reich of the Harvard Medical School in Boston, USA has collated more than 15 thousand ancient human genomes extracted from the remains of individuals who lived and died in Europe and parts of the Middle East during the last ten thousand years. These have been analysed statistically in the context of ‘directional selection’. This is a type of natural selection that occurs when one version of a gene – an allele – confers an extreme form of a trait. If it proves advantageous it rapidly gets passed on to more descendants than do less advantageous alleles, and thus rises in frequency across a population. This differs from other causes of gene frequency changes, such as human migration, population mixing, and random genetic fluctuations that occur in small populations. Well-known examples of directional selection are rapid changes among European Peppered moths, African cichlid fish, Alaskan Sockeye salmon and Big Cats which change over time in response to variations in their habitats. A human example is a genetic variant that maintains the ability to digest the sugar lactose in milk beyond infancy, which enables many modern Europeans to digest milk throughout their lives. The algorithm needed to separate signs of directional selection from other types of genetic change was developed by Ali Akbari, a computational geneticist also at Harvard Medical School. A recent paper by Akbari, Reich and colleagues in the US, Iran, Germany, and Austria (Akbari, A. and 15 others 2026. Ancient DNA reveals pervasive directional selection across West Eurasia. Nature, advance online publication; DOI: 10.1038/s41586-026-10358-1) seems set dramatically to change the research into recent human evolutionary genetics.
Akbari et al. discovered that directional selection has driven the spread or decline of hundreds of gene variants in human populations throughout Western Europe in the last ten millennia. In particular, selection accelerated with the adoption of farming rather than a hunter-gatherer lifestyle. Among the gene variants are those connected with light skin, red hair, risk of celiac disease – linked to gluten in cereals – susceptibility to gout, resistance to leprosy, baldness, rheumatoid arthritis and alcoholism. There are many more (see Figure 3 in the paper): the team identified 479 gene variants affected by directional selection, some that can be explained by changes in lifestyle, others less explicable and yet more that underlie complex traits such as mental illness and cognition. Some of the variants sprang up and were sustained in the population, others rose and then dwindled. The Neolithic began a period of fundamental life style changes in Europe, summed up as a shift from hunting and foraging to farming of cereals and livestock, as early as about 10 ka ago in what is now Türkiye. The pace of genetic changes of this kind reached a peak around the Bronze Age, perhaps because human activities in Europe became more complex then with the mass migration westwards of Yamnaya horse- and wagon-using people from the steppes to dominate Europe
The shift from small wandering bands to living in settlements was a drastic change from a lifestyle that had continued throughout all previous human history. So, it is hardly surprising that there was a major shift in humans’ genetic makeup. But such a change in human labour was not unique to Europe and is known to have occurred on all inhabited continents, with the exception of Australia, at different times during the Holocene. Other regional genetic databases can be analysed in much the same way, once sufficient ancient DNA is collected in Asia, Africa and the Americas. Yet not much is available. The authors comment: ‘A variant that now correlates to household income or years of schooling [remarkably, there are some!] had to have meant something different in the Stone Age. So these results do not mean that Europeans evolved to be smarter or healthier.’ Moreover, the research results in the paper seem likely to be amplified as the data set is so large and complex.
The emergence of the eukaryotes – of which we are a late-entry member – has been debated for quite a while. In 2023 Earth-logs reportedthat a study of ‘biomarker’ organic chemicals in Proterozoic sediments suggests that eukaryotes cannot be traced back further than about 900 Ma ago using such an approach. At about the same time another biomarker study showed signs of a eukaryote presence at around 1050 Ma. Both outcomes seriously contradicted a ‘molecular-clock’ approach based on the DNA of modern members of the Eukarya and estimates of the rate of genetic mutation. That method sought to deduce the time in the past when the last eukaryotic common ancestor (LECA) appeared. It pointed to about 2 Ga ago, i.e. a few hundred million years after the Great Oxygenation Event got underway. Since eukaryote metabolism depends on oxygen, the molecular-clock result seems reasonable. The biomarker evidence does not. But were the Palaeo- and Mesoproterozoic Eras truly ‘boring’? A recent paper by Dietmar Müller and colleagues from the Universities of Sydney and Adelaide, Australia definitely shows that geologically they were far from that (Müller, R.D. et al. 2025. Mid-Proterozoic expansion of passive margins and reduction in volcanic outgassing supported marine oxygenation and eukaryogenesis. Earth and Planetary Science Letters, v. 672; DOI: 10.1016/j.epsl.2025.119683).
Carbon influx (million tons per year) into tectonic plates and into the ocean-atmosphere system from 1800 Ma to present. The colour bands represent: total carbon influx into the atmosphere (mauve); sequestered in tectonic plates (green); net atmospheric influx i.e. total minus carbon sequestered into plates (orange). The widths of the bands show the uncertainties of the calculated masses shown as darker coloured lines.
From 1800 to 800 Ma two supercontinents– Nuna-Columbia and Rodinia – aggregated nearly all existing continental masses, and then broke apart. Continents had collided and then split asunder to drift. So plate tectonics was very active and encompassed the entire planet, as Müller et al’s palaeogeographic animation reveals dramatically. Tectonics behaved in much the same fashion through the succeeding Neoproterozoic and Phanerozoic to build-up then fragment the more familiar supercontinent of Pangaea. Such dynamic events emit magma to form new oceanic lithosphere at oceanic rift systems and arc volcanoes above subduction zones, interspersed with plume-related large igneous provinces and they wax and wane. Inevitably, such partial melting delivered carbon dioxide to the atmosphere. Reaction on land and in the rubbly flanks of spreading ridges between new lithosphere and dissolved CO2 drew down and sequestered some of that gas in the form of solid carbonate minerals. Continental collisions raised the land surface and the pace of weathering, which also acted as a carbon sink. But they also involved metamorphism that released carbon dioxide from limestones involved in the crustal transformation. This protracted and changing tectonic evolution is completely bound up through the rock cycle with geochemical change in the carbon cycle.
From the latest knowledge of the tectonic and other factors behind the accretion and break-up of Nuna and Rodinia, Müller et al. were able to model the changes in the carbon cycle during the ‘boring billion’ and their effects on climate and the chemistry of the oceans. For instance, about 1.46 Ga ago, the total length of continental margins doubled while Nuna broke apart. That would have hugely increased the area of shallow shelf seas where living processes would have been concentrated, including the photosynthetic emission of oxygen. In an evolutionary sense this increased, diversified and separated the ecological niches in which evolution could prosper. It also increased the sequestration of greenhouse gas through reactions on the flanks of a multiplicity of oceanic rift systems, thereby cooling the planet. Translating this into a geochemical model of the changing carbon cycle (see figure) suggests that the rate of carbon addition to the atmosphere (outgassing) halved during the Mesoproterozoic. The carbon cycle and probable global cooling bound up with Nuna’s breakup ended with the start of Rodinia’s aggregation about 1000 Ma ago and the time that biomarkers first indicate the presence of eukaryotes.
Simplified structures of (a) a prokaryote cell; (b) a simple eukaryote animal cell. Plants also contain organelles called chloroplasts
So, did tectonics play a major role in the rise of the Eukarya? Well, of course it did, as much as it was subsequently the changing background to the appearance of the Ediacaran animals and the evolutionary carnival of the Phanerozoic. But did it affect the billion-year delay of ‘eukaryogenesis’ during prolonged availability of the oxygen that such a biological revolution demanded? Possibly not. Lyn Margulis’s hypothesis of the origin of the basic eukaryote cell by a process of ‘endosymbiosis’ is still the best candidate 50 years on. She suggested that such cells were built from various forms of bacteria and archaea successively being engulfed within a cell wall to function together through symbiosis. Compared with prokaryote cells those of the eukaryotes are enormously complex. At each stage the symbionts had to be or become compatible to survive. It is highly unlikely that all components entered the relationship together. Each possible kind of cell assembly was also subject to evolutionary pressures. This clearly was a slow evolutionary process, probably only surviving from stage to stage because of the global presence of a little oxygen. But the eukaryote cell may also have been forced to restart again and again until a stable form emerged.
The colours of human skin, eyes and hair in living people across the world are determined by variants of genes (alleles) found at the same place on a chromosome. Since chromosomes are inherited from both mother and father, an individual may have the same two alleles (homozygous), or one of each (heterozygous). A dominant allele is always expressed, even if a single copy is present. A recessive allele is only expressed if the individual inherits two copies of it. Most characteristics of individuals result from the interaction of multiple genes, rather than a single gene. A commonly cited example is the coloration of eyes. If we had a single gene for eye colour – that of the iris – that had alleles just for blue (recessive or ‘b’) and one for brown (dominant or ‘B) pigmentation, brown-eyed individuals would have one or two ‘B’ alleles (bB or BB), whereas those with blue eyes would have to have two ‘blue’ alleles (bb). But inheritance is more complicated than that: there are people with green, hazel or grey eyes and even left- and right eyes of different colour. Such examples suggest that there are more than two genes affecting human eye colour, and each must have evolved as a result of mutations. Much the same goes for hair and skin coloration.
A group of scientists from the University of Ferrara in Italy have analysed highly detailed ancient DNA in anatomically modern human remains from Russia (Palaeolithic), Sweden (Mesolithic) and Croatia (Neolithic) to tease out the complexities of pigmentation inheritance. Then they applied a statistical approach learned from that study to predict the likely skin-, eye- and hair pigmentation in 348 less detailed genomes of ancient individuals whose remains date back to 45 Ma ( Silvia Perretti et al, 2025. Inference of human pigmentation from ancient DNA by genotype likelihood. Proceedings of the National Academy of Science, v. 122, article e2502158122; DOI: 10.1073/pnas.2502158122).
An artist’s impression of a Mesolithic woman from southern Denmark (credit: Tom Bjorklund)
All the hunter-gatherer Palaeolithic individuals (12 samples between 45 and 13 ka old) bar one, showed clear signs of dark pigmentation in skin, eyes and hair – the outlier from Russia was probably lighter. Those from the Mesolithic (14 to 4 ka) showed that 11 out of 35 had a light eye colour (Northern Europe, France, and Serbia), but most retained the dark skin and hair expected in descendants of migrants from Africa. Only one 12 ka hunter-gatherer from Sweden had inferred blue eyes, blonde hair, and light skin. The retention of dark pigmentation by European hunter-gatherers who migrated there from Africa has been noted before, using DNA from Mesolithic human remains and in one case from birch resin chewed by a Mesolithic woman. This called into question the hypothesis that high levels of melatonin in skin, which protects indigenous people in Africa from cancers, would result in their producing insufficient vitamin D for good health. That notion supposed that out-of-Africa migrants would quickly evolve paler skin coloration at higher latitudes. It is now known that diets rich in meat, nuts and fungi – staple for hunter-gatherers – provide sufficient vitamin-D for health at high latitudes. A more recent hypothesis is that pale skins may have evolved only after the widespread Neolithic adoption of farming when people came to rely on a diet dominated by cereals that are a poor source of vitamin-D.
However, 132 Neolithic farmers (10 to 4 ka ago) individuals studied by Perretti et al. showed increased diversity in pigmentation, with more frequent light skin tones, yet dark individuals persisted, particularly in southern and eastern Europe. Hair and eye colour showed considerable variability, the earliest sign of red hair showing up in Turkey. Even Copper- and Bronze Age samples ( 113 from 7 to 3 ka) and those from Iron Age Europeans (25 from 3 to 1.7 ka ago) still indicate common retention of dark skin, eyes and hair, although the proportion of lighter pigmentation increased in some regions of Europe. Other analyses of ancient DNA have shown that the Palaeo- and Mesolithic populations of Europe were quickly outnumbered by influx of early farmers, probably from the Anatolian region of modern Turkey, during the Neolithic. The farming lifestyle seems likely to have allowed the numbers of those who practised it to rise beyond the natural environment’s ‘carrying capacity’ for hunter-gatherers. The former inhabitants of Europe may simply have been genetically absorbed within the growing population of farmers. Much the same absorption of earlier groups seems to have happened with the westward migration from the Ukrainian and Russia steppes of the Yamnaya people and culture, culminating in the start of the European Bronze Age that reached western Europe around 2.1 ka, The Yamnaya introduced metal culture, horse-drawn wheeled vehicles and possibly Indo-European language.
So the novel probabilistic approach to ancient DNA by Perretti et al. also casts doubt on the diet-based evolution of light pigmentation at high latitudes. Instead, pulses of large population movements and thus changes in European population genetics probably account for the persistence of abundant evidence for dark pigmentation throughout Europe until historic times. The ‘lightening’ of Europeans’ physiognomy seems to have been vastly more complex than previously believed. Early Europe seems to have been almost bewilderingly diverse, which make a complete mockery of modern chauvinism and racism. The present European genetic ‘melting pot’ is surprisingly similar to that of Europe’s ancient past.
The molecules that make up all living matter are almost entirely (~98 %) made from the elements Carbon, Hydrogen, Oxygen, Nitrogen and Phosphorus (CHONP) in order of their biological importance. All have low atomic numbers, respectively 6th, 1st, 8th, 7th and 15th in the Periodic Table. Of the 98 elements found in nature, about 7 occur only because they form in the decay schemes of radioactive isotopes. Only the first 83 (up to Bismuth) are likely to be around ‘for ever’; the fifteen heavier than that are made up exclusively of unstable isotopes that will eventually disappear, albeit billions of years from now. There are other oddities that mean that the 92 widely accepted to be naturally occurring is not strictly correct. That CHONP are so biologically important stems partly from their abundances in the inorganic world and also because of the ease with which they chemically combine together. But they are not the only ones that are essential.
About 20 to 25% of the other elements are also literally vital, even though many are rare. Most of the rest are inessential except in vanishingly small amounts that do no damage, and may or may not be beneficial. However some are highly toxic. Any element can produce negative biological outcomes if above certain levels. Likewise, deficiencies can result in ill thrift and event death. For the majority of elements, biologists have established concentrations that define deficiency and toxic excess. The World Health Organisation has charted the maximum safe levels of elements in drinking water in milligrams per litre. In this regard, the lowest safe level is for thallium (Tl) and mercury (Hg) at 0.002 mg l-1.Other highly toxic elements are cadmium (Cd) (0.003 mg l-1), then arsenic (As) and lead (Pb) (0.01 mg l-1) that ‘everyone knows’ are elements to avoid like the plague. In nature lead is very rarely at levels that are unsafe because it is insoluble, but arsenic is soluble under reducing conditions and is currently responsible for a pandemic of related ailments, especially in the Gangetic plains of India and Bangladesh and similar environments worldwide.
Biological evolution has been influenced since life appeared by the availability, generally in water, of both essential and toxic elements. In 2020 Earth-logs summarised a paper about modern oxygen-free springs in Chile in which photosynthetic purple sulfur bacteria form thick microbial mats. The springs contain levels of arsenic that vary from high in winter to low in summer. This phenomenon can only be explained by some process that removes arsenic from solution in summer but not in winter. The purple-bacteria’s photosynthesis uses electrons donated by sulfur, iron-2 and hydrogen – the spring water is highly reducing so they thrive in it. In such a simple environment this suggested a reasonable explanation: the bacteria use arsenic too. In fact they contain a gene (aio) that encodes for such an eventuality. The authors suggested that purple sulfur bacteria may well have evolved before the Great Oxygenation Event (GOE). They reasoned that in an oxygen-free world arsenic, as well as Fe2+ would be readily available in water that was in a reducing state, whereas oxidising conditions after the GOE would suppress both: iron-2 would be precipitated as insoluble iron-3 oxides that in turn efficiently absorb arsenic (see: Arsenic hazard on a global scale, May 2020).
Colour photograph and CT scans of Palaeoproterozoic discoidal fossils from the Francevillian Series in Gabon. (Credit: El Albani et al. 2010; Fig. 4).
A group of geoscientists from France, the UK, Switzerland and Austria have investigated the paradox of probably high arsenic levels before the GOE and the origin and evolution of life during the Archaean (El Khoury et al. 2025. A battle against arsenic toxicity by Earth’s earliest complex life forms. Nature Communications, v. 16, article 4388; DOI: 10.1038/s41467-025-59760-9). Note that the main, direct evidence for Archaean life are fossilized microbial mats known as stromatolites, some palaeobiologists reckoning they were formed by oxygenic photosynthesising cyanobacteria others favouring the purple sulfur bacteria (above). The purple sulfur bacteria in Chile and other living prokaryotes that tolerate and even use arsenic in their metabolism clearly evolved that potential plus necessary chemical defence mechanisms, probably when arsenic was more available in the anoxic period before the GOE. Anna El Khoury and her colleagues sought to establish whether or not eukaryotes evolved similar defences by investigating the earliest-known examples; the 2.1 Ma old Francevillian biota of Gabon that post-dates the GOE. They are found in black shales, look like tiny fried eggs and are associated with clear signs of burrowing. The shales contain steranes that are breakdown products of steroids, which are unique to eukaryotes.
The fossils have been preserved by precipitation of pyrite (Fe2S) granules under highly reducing conditions. Curiously, the cores of the pyrite granules in the fossils are rich in arsenic, yet pyrite grains in the host sediments have much lower As concentrations. The latter suggest that seawater 2.1 Ma ago held little dissolved arsenic as a result of its containing oxygen. The authors interpret the apparently biogenic pyrite’s arsenic cores as evidence of the organism having sequestered As into specialized compartments in their bodies: their ancestors must have evolved this efficient means of coping with significant arsenic stress before the GOE. It served them well in the highly reducing conditions of black shale sedimentation. Seemingly, some modern eukaryotes retain an analogue of a prokaryote As detoxification gene.
False-colour electron microscope image of a shocked grain of zircon recovered from the Stac Fada Member. The red and pink material is a high-pressure polymorph of zircon, arranged in shock lamellae. Zircon is rendered in cyan, some of which is in granulated form. Credit: Kirkland et al. 2025, Fig 2C
Judging by its content of shards and spherules made of murky green glass, one of the lowest units in the Torridonian continental sediments of NW Scotland had long been regarded as simply red sandstone that contained volcanic debris. This Stac Fada Member was thus celebrated as the only sign of a volcanic contribution to a vast thickness (up to 2.5 km) of Neoproterozoic lake and fluviatile sediments. Current flow indicators suggested that the Torridonian was laid down by large alluvial fans derived by erosion of much older crystalline basement far to what is today the west. That is, the Archaean core of the ancient continent of Laurentia, now the other side of the North Atlantic. In 2002 more sophisticated sedimentological and geochemical analysis of the Stac Fada Member revealed a surprise: it contains anomalously elevated platinum-group elements, quartz grains that show signs of shock and otherworldly chromium isotope concentrations. The 10 m thick bed is made from ejecta, perhaps from a nearby impact crater to the WNW concluded from brittle fractures that may have been produced by the impact. Some idea of its age was suggested by Ar-Ar dating of feldspar crystals (~1200 Ma) believed to have formed authigenically in the hot debris. Being the only decent impactite known in Britain, it continues to attract attention.
A group of geoscientists from Western Australia, NASA and the UK, independent of the original discoverers, have now added new insights ( Kirkland, C.L. and 12 others 2025. A one-billion-year old Scottish meteorite impact. Geology, v. 53, early online publication; DOI: 10.1130/G53121.1). They dated shocked zircon grains using U-Pb analyses at 990 ± 22 Ma; some 200 Ma younger than the previously dated, authigenic feldspars. Detrital feldspar grains in the Stac Fada Member yield Rb-Sr radiometric ages of 1735 and 1675, that are compatible with Palaeoproterozoic granites in the underlying Lewisian Gneiss Complex.
Photomicrograph of Bicellum brazieiri: scale bar = 10μm; arrows point to dark spots that may be cell nuclei (credit: Charles Wellman, Sheffield University)
In a separate publication (Kirkland, C.L et al 2025. 1 billion years ago, a meteorite struck Scotland and influenced life on Earth. The Conversation, 29 April 2025) three of the authors take things a little further, as their title suggests. In this Conversation piece they ponder, perhaps unwarily, on the spatial and temporal association of the indubitable impact with remarkably well-preserved spherical fossils found in Torridonian lake-bed sediments (Bicellum brasieri, reported in Earth-logs in May 2021), which are the earliest-known holozoan animal ancestors. The Torridonian phosphatic concretions in which these important fossils were found at a different locality are roughly 40 Ma younger than the Stac Fada impactite. The authors of the Conversation article appeal to the residual thermal effect of the impact as a possible driver for the appearance of these holozoan organisms. Whether a residual thermal anomaly would last long enough for them to evolve to this biological status would depend on the magnitude of the impact, of which we know nothing. Eukaryote fossils are known from at least 650 Ma older sedimentary rocks in northern China and perhaps as far back as 2.2 Ga in a soil that formed in the Palaeoproterozoic of South Africa. Both the Torridonian organism and impactite were found in a small area of fascinating geology that has been studied continuously in minute detail since Victorian times, and visited by most living British geologists during their undergraduate days. Ideas will change as curiosity draws geologists and palaeobiologists to less-well studied sites of Proterozoic antiquity, quite possibly in northern China.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
A consortium of geoscientists from Australia, Britain and France, led by Andrew Merdith of the University of Adelaide examines the likely climate cooling mechanisms that may have set off the two great ‘icehouse’ intervals in the last 541 Ma (Merdith, A.S. et al. 2025. Phanerozoic icehouse climates as the result of multiple solid-Earth cooling mechanisms. Science Advances, v. 11, article eadm9798: DOI: 10.1126/sciadv.adm9798). They consider the first to be the global cooling that began in the latter part of the Devonian culminating in the Carboniferous-Permian icehouse. The second is the Cenozoic global cooling to form the permanent Antarctic ice cap around 34 Ma and culminated in cyclical ice ages on the northern continents after 2.4 Ma during the Pleistocene. They dismiss the 40 Ma long, late Ordovician to early Silurian glaciation that left its imprint on North Africa and South America – then combined in the Gondwana supercontinent. The data about two of the parameters used in their model – the degree of early colonisation of the continents by plants and their influence on terrestrial weathering are uncertain in that protracted event. Yet the Hirnantian glaciation reached 20°S at its maximum extent in the Late Ordovician around 444 Ma to cover about a third of Gondwana: it was larger than the present Antarctic ice cap. For that reason, their study spans only Devonian and later times.
Fluctuation in evidence for the extent of glacial conditions since the Devonian: the ‘ice line’ is grey. The count of glacial proxy occurrences in each 10° of latitude through time is shown in the colour key. Credit: Merdith et al., Fig 2A.
Merdith et al. rely on four climatic proxies. The first of these comprises indicators of cold climates, such as glacial dropstones, tillites and evidence in sedimentary rocks of crystals of hydrated calcium carbonate (ikaite – CaCO3.6H2O) that bizarrely forms only at around 0°C . From such occurrences it is possible to define an ‘ice line’ linking different latitudes through geological time. Then there are estimates of global average surface temperature; low-latitude sea surface temperature; and estimates of atmospheric CO2. The ‘ice-line’ data records an additional, long period of glaciation in the Jurassic and early Cretaceous, but evidence does not extend to latitudes lower than 60°. It is regarded by Merdith et al. as an episode of ‘cooling’ rather than an ‘icehouse’. Their model assesses sources and sinks of CO2 since the Devonian Period.
The main natural source of the principal greenhouse gas CO2 is degassing through volcanism expelled from the mantle and breakdown of carbonate rock in subducted lithosphere. Natural sequestration of carbon involves weathering of exposed rock that releases dissolved CO2 and ions of calcium and magnesium. A recently compiled set of plate reconstructions that chart the waxing and waning of tectonics since the Devonian Period allows them to model the tectonically driven release of carbon over time, with time scales on the order of tens to hundreds of Ma. The familiar Milanković forcing cycles on the order of tens to hundreds of ka are thus of no significance in Merdith et al.’s broader conception of icehouse episodes Their modelling shows high degassing during the Cretaceous, modern levels during the late Palaeozoic and early Mesozoic, and low emissions during the Devonian. The model also suggests that cooling stemmed from variations in the positions and configuration of continents over time. Another crucial factor is the tempo of exposure of rocks that are most prone to weathering. The most important are rocks of the ocean lithosphere incorporated into the continents to form ophiolite masses. The release of soluble products of weathering into ocean basins through time acts as a fluctuating means of ‘fertilising’ so that more carbon can be sequestered in deep sediments in the form of organisms’ unoxidised tissue and hard parts made of calcium carbonates and phosphates. Less silicate weathering results in a boost to atmospheric CO2.
Only two long, true icehouse episodes emerge from the empirical proxy data, expressed by the ‘ice-line’ plots. Restricting the modelling to single global processes that might be expected to influence degassing or carbon sequestration produces no good fits to the climatic proxy data. Running the model with all the drivers “off” produces more or less continuous icehouse conditions since the Devonian. The model’s climate-related outputs thus imply that many complex processes working together in syncopation may have driven the gross climate vagaries over the last 400 Ma or so. A planet of Earth’s size without such complexity would throughout that period have had a high-CO2 warm climate. According to Andrew Merdith its fluctuation from greenhouse to icehouse conditions in the late Palaeozoic and the Cenozoic were probably due to “coincidental combination of very low rates of global volcanism, and highly dispersed continents with big mountains, which allow for lots of global rainfall and therefore amplify reactions that remove carbon from the atmosphere”.
Geological history is, almost by definition, somewhat rambling. So, despite despite the large investment in seeking a computed explanation of data drawn from the record, the outcome reflects that in a less than coherent account. To state that many complex processes working at once may have driven climate vagaries over the last 400 Ma or so, is hardly a major advance: palaeoclimatologists have said more or less the same for a couple of decades or more, but have mainly proposed single driving mechanisms. One aspect of Merdith et al.’s results seems to be of particular interest. ‘Icehouse’ conditions seem to be rare events interspersed with broader ice-free periods. We evolved within the mammal-dominated ecosystems on the continents during the latest of these anomalous climatic episodes. And we and those ecosystems now rely on a cool world. As the supervisor of the project commented, ‘Over its long history, the Earth likes it hot, but our human society does not’.
Readers may like to venture into how some philosophers of science deal with a far bigger question; ‘Is intelligent life a rare, chance event throughout the universe?’ That is, might we be alone in the cosmos? In the same issue of Science Advances is a paper centred on just such questions (Mills, D.B. et al. 2025. A reassessment of the “hard-steps” model for the evolution of intelligent life. Science Advances, v. 11, article eads5698; DOI: 10.1126/sciadv.ads5698). It stems from cosmologist Brandon Carter’s ‘Anthropic Principle’ first developed at Nicolas Copernicus’s 500th birthday celebrations in 1973. This has since been much debated by scientists and philosophers – a gross understatement as it knocks the spots off the Drake Equation. To take the edge off what seems to be a daunting task, Mills et al. consider a corollary of the Anthropic Principle, the ‘hard steps model’. That, in a nutshell, postulates that the origin of humanity and its ability to ponder on observations of the universe required a successful evolutionary passage through a number of hard steps. It predicts that such intelligence is ‘exceedingly rare’ in the universe. Icehouse conditions are respectable candidates for evolutionary ‘hard steps’, and in the history of Earth there have been five of them.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Debates around the origin of Earth’s life and what the first organism was like resemble the mythical search for the Holy Grail. Chivalric romanticists of the late 12th and early 13th centuries were pretty clear about the Grail – some kind of receptacle connected either with the Last Supper or Christ’s crucifixion – but never found it. Two big quests that engage modern science centre on how the chemical building blocks of the earliest cells arose and the last universal common ancestor (LUCA) of all living things. Like the Grail’s location, neither is likely to be fully resolved because they can only be sought in a very roundabout way: both verge on the imaginary. The fossil record is limited to organisms that left skeletal remains, traces of their former presence, and a few degraded organic molecules. The further back in geological time the more sedimentary rock has either been removed by erosion or fundamentally changed at high temperatures and pressures. Both great conundrums can only be addressed by trying to reconstruct processes and organisms that occurred or existed more than 4 billion years ago.
Artistic impression of the early Earth dominated by oceans (Credit: Sci-news.com)
In the 1950s Harold Urey of the University of Chicago and his student Stanley Miller mixed water, methane, ammonia and hydrogen sulfide in lab glassware, heated it up and passed electrical discharges through it. They believed the simple set-up crudely mimicked Hadean conditions at the Earth surface. They were successful in generating more complex organic chemicals than their starting materials, though the early atmosphere and oceans are now considered to have been chemically quite different. Such a ‘Frankenstein’ approach has been repeated since with more success (see Earth-logs April 2024), creating 10 of the 20 amino acids plus the peptide bonds that link them up to make all known proteins, and even amphiphiles, the likely founders of cell walls. The latest attempt has been made by Spanish scientists at the Andalusian Earth Sciences Institute, the Universities of Valladolid and Cadiz, and the International Physics Centre in San Sebastian (Jenewein, C. et al 2024. Concomitant formation of protocells and prebiotic compounds under a plausible early Earth atmosphere. Proceedings of the National Academy of Sciences, v. 122, article 413816122; DOI: 10.1073/pnas.241381612).
Biomorphs formed by polymerisation of HCN (Credit: Jenewein, C. et al 2024, Figure 2)
Jenewein and colleagues claim to have created cell-like structures, or ‘biomorphs’ at nanometre- and micrometre scale – spheres and polyp-like bodies – from a more plausible atmosphere of CO2 , H2O, and N2. These ‘protocells’ seem to have formed from minutely thin (150 to 3000 nanometres) polymer films built from hydrogen cyanide that grew on the surface of the reaction chamber as electric discharges and UV light generated HCN and more complex ‘prebiotic’ chemicals. Apparently, these films were catalysed by SiO2 (silica) molecules from the glass reactor. Note: In the Hadean breakdown of olivine to serpentinite as sea water reacted with ultramafic lavas would have released abundant silica. Serpentinisation also generates hydrogen. Intimate release of gas formed bubbles to create the spherical and polyp-like ‘protocells’. The authors imagine the Hadean global ocean permanently teeming with such microscopic receptacles. Such a veritable ‘primordial soup’ would be able to isolate other small molecules, such as amino acids, oligopeptides, nucleobases, and fatty acids, to generate more complex organic molecules in micro-reactors en route to the kind of complex, self-sustaining systems we know as life.
So, is it possible to make a reasonable stab at what that first kind of life may have been? It was without doubt single celled. To reproduce it must have carried a genetic code enshrined in DNA, which is unique not only to all species, but to individuals. The key to tracking down LUCA is that it represents the point at which the evolutionary trees of the fundamental domains of modern life life – eukarya (including animals, plants and fungi), bacteria, and archaea – converge to a single evolutionary stem. There is little point in using fossils to resolve this issue because only multicelled life leaves tangible traces, and the first of those was found in 2,100 Ma old sediments in Gabon (see: The earliest multicelled life; July 2010). The key is using AI to compare the genetic sequences of the hugely diverse modern biosphere. Modern molecular phylogenetics and computing power can discern from their similarities and differences the relative order in which various species and broader groups split from others. It can also trace the origins of specific genes that provides clues about earlier genetic associations. Given a rate of mutation the modern differences provide estimates of when each branching occurred. The most recent genetic delving has been achieved by a consortium based at various institutions in Britain, the Netherlands, Hungary and Japan (Moody, E.R.R. and 18 others 2024. The nature of the last universal common ancestor and its impact on the early Earth system. Nature Ecology & Evolution, v.8, pages 1654–1666; DOI: 10.1038/s41559-024-02461-1).
Moody et al have pushed back the estimated age of LUCA to halfway through the Hadean, between 4.09 to 4.33 billion years (Ga), well beyond the geologically known age of the earliest traces of life (3.5 Ga). That age for LUCA in itself is quite astonishing: it could have been only a couple of hundred million years after the Moon-forming interplanetary collision. Moreover, they have estimated that Darwin’s Ur-organism had a genome of around 2 million base pairs that encoded about 2600 proteins: roughly comparable to living species of bacteria and archaea, and thus probably quite advanced in evolutionary terms. The gene types probably carried by LUCA suggest that it may have been an anaerobic acetogen; i.e. an organism whose metabolism generated acetate (CH3COO−) ions. Acetogens may produce their own food as autotrophs, or metabolise other organisms (heterotrophs). If LUCA was a heterotroph, then it must have subsisted in an ecosystem together with autotrophs which it consumed, possibly by fermentation. To function it also required hydrogen that can be supplied by the breakdown of ultramafic rocks to serpentinites, which tallies with the likely ocean-world with ultramafic igneous crust of the Hadean (see the earlier paragraphs about protocells). If an autotroph, LUCA would have had an abundance of CO2 and H2 to sustain it, and may have provided food for heterotrophs in the early ecosystem. The most remarkable possibility discerned by Moody et al is that LUCA may have had a kind of immune system to stave off viral infection.
The carbon cycle on the Hadean Earth (Credit: Moody et al. 2024; Figure 3e)
The Hadean environment was vastly different to that of modern times: a waterworld seething with volcanism; no continents; a target for errant asteroids and comets; more rapidly spinning with a 12 hour day; a much closer Moon and thus far bigger tides. The genetic template for the biosphere of the following four billion years was laid down then. LUCA and its companions may well have been unique to the Earth, as are their descendants. It is hard to believe that other worlds with the potential for life, even those in the solar system, could have followed a similar biogeochemical course. They may have life, but probably not as we know it . . .
Refugees from the Middle East migrating through Slovenia in 2015. Credit: Britannica
During the Pliocene (5.3 to 2.7 Ma) there evolved a network of various hominins, with their remains scattered across both the northern and southern parts of that continent. The earliest, though somewhat disputed hominin fossil Sahelanthropus tchadensis hails from northern Chad and lived around 7 Ma ago, during the late Miocene, as did a similarly disputed creature from Kenya Orrorin tugenensis (~5.8 Ma). The two were geographically separated by 1500 km, what is now the Sahara desert and the East African Rift System. The suggestion from mtDNA evidence that humans and chimpanzees had a common ancestor, the uncertainty about when it lived (between 13 to 5 Ma) and what it may have looked like, let alone where it lived, makes the notion debateable. There is even a possibility that the common ancestor of humans and the other anthropoid apes may have been European. Its descendants could well have crossed to North Africa when the Mediterranean Sea had been evaporated away to form the thick salt deposits that now lie beneath it: what could be termed the ‘Into Africa’ hypothesis. The better known Pliocene hominins were also widely distributed in the east and south of the African continent. Wandering around was clearly a hominin predilection from their outset. The same can be said about humans in the general sense (genus Homo) during the Early Pleistocene when some of them left Africa for Eurasia. Artifacts dated at 2.1 Ma have been found on the Loess Plateau of western China, and Georgia hosts the earliest human remains known from Eurasia. Since them H. antecessor, heidelbergensis, Neanderthals and Denisovans roamed Eurasia. Then, after about 130 ka, anatomically modern humans progressively populated all continents, except Antarctica, to their geographic extremities and from sea level to 4 km above it.
There is a popular view that curiosity and exploration are endemic and perhaps unique to the human line: ‘It’s in our genes’. But even plants migrate, as do all animal species. So it is best to be wary of a kind of hominin exceptionalism or superior motive force. Before settled agriculture, simply diffusion of populations in search of sustenance could have achieved the enormous migrations undertaken by all hominins: biological resources move and hunter gatherers follow them. The first migration of Homo erectus from Africa to northern China by way of Georgia seems to taken 200 ka at most and covered about ten thousand kilometres: on average a speed of only 50 m per year! That achievement and many others before and later were interwoven with the evolution of brain size, cognitive ability, means of communication and culture. But what were the ultimate drivers? Two recent papers in the journal Nature Communications make empirically-based cases for natural forces driving the movement of people and changes in demography.
The first considers hominin dispersal in the Palaearctic biogeographic realm: the largest of eight originally proposed by Alfred Russel Wallace in the late 19th century that encompasses the whole of Eurasia and North Africa (Zan, J. et al. 2024. Mid-Pleistocene aridity and landscape shifts promoted Palearctic hominin dispersals. Nature Communications, v. 15, article 10279; DOI: 10.1038/s41467-024-54767-0). The Palearctic comprises a wide range of ecosystems: arid to wet, tropical to arctic. After 2 Ma ago, hominins moved to all its parts several times. The approach followed by Zan et al. is to assess the 3.6 Ma record of the thick deposits of dust carried by the perpetual westerly winds that cross Central Asia. This gave rise to the huge (635,000 km2) Loess Plateau. At least 17 separate soil layers in the loess have yielded artefacts during the last 2.1 Ma. The authors radiocarbon dated the successive layers of loess in Tajikistan (286 samples) and the Tarim Basin (244 samples) as precisely as possible, achieving time resolutions of 5 to 10 ka and 10 to 20 ka respectively. To judge variations in climate in these area they also measured the carbon isotopic proportions in organic materials preserved within the layers. Another climate-linked metric that Zan et al. is a time series showing the development of river terraces across Eurasia derived from the earlier work of many geomorphologists. The results from those studies are linked to variations through time in the numbers of archaeological sites across Eurasia that have yielded hominin fossils, stone tools and signs of tool manufacture, many of which have been dated accurately.
The authors use sophisticated statistics to find correlations between times of climatic change and the signs of hominin occupation. Episodes of desertification in Palaearctic Eurasia clearly hindered hominins’ spreading across the continent either from west to east of vice versa. But there were distinct, periodic windows of climatic opportunity for that to happen that coincide with interglacial episodes, whose frequency changed at the Mid Pleistocene Transition (MPT) from about 41 ka to roughly every 100 ka. That was suggested in 2021 to have arisen from an increased roughness of the rock surface over which the great ice sheets of the Northern Hemisphere moved. This suppressed the pace of ice movement so that the 41 ka changes in the tilt of the Earth’s rotational axis could no longer drive climate change during the later Pleistocene, despite the fact that the same astronomical influence continued. The succeeding ~100 ka pulsation may or may not have been paced by the very much weaker influence of Earth changing orbital eccentricity. Whichever, after the MPT climate changes became much more extreme, making human dispersal in the Palearctic realm more problematic. Rather than hominin’s evolution driving them to a ‘Manifest Destiny’ of dominating the world vastly larger and wider inorganic forces corralled and released them so that, eventually, they did.
Much the same conclusion, it seems to me, emerges from a second study that covers the period since ~ 9 ka ago when anatomically modern humans transitioned from a globally dominant hunter-gatherer culture to one of ‘managing’ and dominating ecosystems, physical resources and ultimately the planet itself. (Wirtz, K.W et al. 2024. Multicentennial cycles in continental demography synchronous with solar activity and climate stability. Nature Communications, v. 15, article 10248; DOI: 10.1038/s41467-024-54474-w). Like Zan et al., Kai Wirtz and colleagues from Germany, Ukraine and Ireland base their findings on a vast accumulated number (~180,000) of radiocarbon dates from Holocene archaeological sites from all inhabited continents. The greatest number (>90,000) are from Europe. The authors applied statistical methods to judge human population variations since 11.7 ka in each continental area. Known sites are probably significantly outweighed by signs of human presence that remain hidden, and the diligence of surveys varies from country to country and continent to continent: Britain, the Netherlands and Southern Scandinavia are by far the best surveyed. Given those caveats, clearly this approach gives only a blurred estimate of population dynamics during the Holocene. Nonetheless the data are very interesting.
The changes in population growth rates show distinct cyclicity during the Holocene, which Wirtz et al. suggest are signs of booms and busts in population on all six continents. Matching these records against a large number of climatic time series reveals a correlation. Their chosen metric is variation in solar irradiance: the power per unit area received from the Sun. That has been directly monitored only over a couple of centuries. But ice cores and tree rings contain proxies for solar irradiance in the proportions of the radioactive isotopes 10Be and 14C contained in them respectively. Both are produced by the solar wind of high-energy charged particles (electrons, protons and helium nuclei or alpha particles) penetrating the upper atmosphere. The two isotopes have half-lives long enough for them to remain undecayed and thus detectable for tens of thousand years. Both ice cores and tree rings have decadal to annual time resolutions. Wirtz et al. find that their crude estimates of booms and busts in human populations during the Holocene seem closely to match variations in solar activity measured in this way. Climate stability favours successful subsistence and thus growth in populations. Variable climatic conditions seem to induce subsistence failures and increase mortality, probably through malnutrition.
A nice dialectic clearly emerges from these studies. ‘Boom and bust’ as regards populations in millennial and centennial to decadal terms stem from climate variations. Such cyclical change thus repeatedly hones natural selection among the survivors, both genetically and culturally, increasing their general fitness to their surroundings. Karl Marx and Friedrich Engels would have devoured these data avidly had they emerged in the 19th century. I’m sure they would have suggested from the evidence that something could go badly wrong – negation of negation, if readers care to explore that dialectical law further . . . And indeed that is happening. Humans made ecologically very fit indeed in surviving natural pressures are now stoking up a major climatic hiccup, or rather the culture and institutions that humans have evolved are doing that.
For the last forty thousand years anatomically modern humans have been the only primates living on Planet Earth with a sophisticated culture; i.e. using tools, fire, language, art etcetera. Since Homo sapiens emerged some 300 ka ago, they joined at least two other groups of humans – Neanderthals and Denisovans – and not only shared Eurasia with them, but interbred as well. In fact no hominin group has been truly alone since Pliocene times, which began 5.3 Ma ago. Sometimes up to half a dozen species occupied the habitable areas of Africa. Yet we can never be sure whether or not they bumped into one another. Dates for fossils are generally imprecise; give or take a few thousand years. The evidence is merely that sedimentary strata of roughly the same age in various places have yielded fossils of several hominins, but that co-occupation has never been proved in a single stratum in the same place: until now.
Footprints from Koobi Fora: left – right foot of H. erectus; right – left foot of Paranthropus boisei. Credit: Kevin Hatala. Chatham University
The Koobi Fora area near modern Lake Turkana has been an important, go-to site, courtesy of the Leakey palaeoanthropology dynasty (Louis and Mary, their son and daughter-in-law Richard and Meave, and granddaughter Louise). They discovered five hominin species there dating from 4.2 to 1.4 Ma. So there was a chance that this rich area might prove that two of the species were close neighbours in both space and time. In 2021 Kenyan members of the Turkana Basin Institute based in Nairobi spotted a trackway of human footprints on a bedding surface of sediments that had been deposited about 1.5 Ma ago. Reminiscent of the famous, 2 million years older Laetoli trackway of Australopithecus afarensis in Tanzania, that at Koobi Fora is scientifically just as exciting for it shows footprints of two hominin species Homo erectus and Paranthropus boisei who had walked through wet mud a few centimetres below the surface of Lake Turkana’s ancient predecessor (Hatala, K.G. and 13 others, 2024. Footprint evidence for locomotive diversity and shared habitats among early Pleistocene hominins. Science, v. 386, p. 1004-1010; DOI: 10.1126/science.ado5275). The trackway is littered with the footprints of large birds and contains evidence of zebra.
One set of prints attributed to H. erectus suggest the heels struck the surface first, then the feet rolled forwards before pushing off with the soles: little different from our own, unshod footprints in mud. They are attributed to H. erectus. The others also show a bipedal gait, but different locomotion. The feet that made them were significantly flatter than ours and had a big toe angled away from the smaller toes. They are so different that no close human relative could have made them. The local fossil record includes paranthropoids (Paranthropus boisei), whose fossil foot bones suggest an individual of that speciesmade those prints. It also turns out that a similar, dual walkers’ pattern was found 40 km away in lake sediments of roughly the same age. The two species cohabited the same terrain for a substantial period of time. As regards the Koobi Fora trackway, it seems the two hominins plodded through the mud only a few hours apart at most: they were neighbours.
Artists’ reconstructions of: left – H. erectus; right – Paranthropus boisei. Credits: Yale University, Roman Yevseyev respectively
From their respective anatomies they were very different. Homo erectus was, apart from having massive brow ridges, similar to us. Paranthropus boisei had huge jaws and facial muscles attached to a bony skull crest. So how did they get along? The first was probably omnivorous and actively hunted or scavenged meaty prey: a bifacial axe-wielding hunter-gatherer. Paranthropoids seem to have sought and eaten only vegetable victuals, and some sites preserve bone digging sticks. They were not in competition for foodstuffs and there was no reason for mutual intolerance. Yet they were physically so different that intimate social relations were pretty unlikely. Also their brain sizes were very different, that of P. Boisei’s being far smaller than that of H. erectus , which may not have encouraged intellectual discourse. Both persist in the fossil record for a million years or more. Modern humans, Neanderthals and Denisovans, as we know, sometimes got along swimmingly, possibly because they were cognitively very similar and not so different physically.
Since many hominin fossils are associated with riverine and lake-side environments, it is surprising that more trackways than those of Laetoli and Koobi Fora have been found. Perhaps that is because palaeoanthropologists are generally bent on finding bones and tools! Yet trackways show in a very graphic way how animals behave and interrelate with their environment, for example dinosaurs. Now anthropologists have learned how to spot footprint trace fossils that will change, and enrich the human story.
In March 1989 an asteroid half a kilometre across passed within 500 km of the Earth at a speed of 20 km s-1. Making some assumptions about its density, the kinetic energy of this near miss would have been around 4 x 1019 J: a million times more than Earth’s annual heat production and humanity’s annual energy use; and about half the power of detonating every thermonuclear device ever assembled. Had that small asteroid struck the Earth all this energy would have been delivered in a variety of forms to the Earth System in little more than a second – the time it would take to pass through the atmosphere. The founder of “astrogeology” and NASA’s principal geological advisor for the Apollo programme, the late Eugene Shoemaker, likened the scenario to a ‘small hill falling out of the sky’. (Read a summary of what would happen during such an asteroid strike). But that would have been dwarfed by the 10 to 15 km impactor that resulted in the ~200 km wide Chicxulub crater and the K-Pg mass extinction 66 Ma ago. Evidence has been assembled for Earth having been struck during the Archaean around 3.6 billion years (Ga) ago by an asteroid 200 to 500 times larger: more like four Mount Everests ‘falling out of the sky’ (Drabon, N. et al. 2024. Effect of a giant meteorite impact on Paleoarchean surface environments and life. Proceedings of the National Academy of Sciences, v. 121, article e2408721121; DOI: 10.1073/pnas.2408721121
Impact debris layer in the Palaeoarchaean Barberton greenstone belt of South Africa, which contains altered glass spherules and fragments of older carbonaceous cherts. (Credit: Credit: Drabon, N. et al., Appendix Fig S2B)
In fact the Palaeoarchaean Era (3600 to 3200 Ma) was a time of multiple large impacts. Yet their recognition stems not from tangible craters but strata that contain once glassy spherules, condensed from vaporised rock, interbedded with sediments of Palaeoarchaean ‘greenstone belts’ in Australia and South Africa (see: Evidence builds for major impacts in Early Archaean; August 2002, and Impacts in the early Archaean; April 2014), some of which contain unearthly proportions of different chromium isotopes (see: Chromium isotopes and Archaean impacts; March 2003). Compared with the global few millimetres of spherules at the K-Pg boundary, the Barberton greenstone belt contains eight such beds up to 1.3 m thick in its 3.6 to 3.3 Ga stratigraphy. The thickest of these beds (S2) formed by an impact at around 3.26 Ga by an asteroid estimated to have had a mass 50 to 200 times that of the K-Pg impactor.
Above the S2 bed are carbonaceous cherts that contain carbon-isotope evidence of a boom in single-celled organisms with a metabolism that depended on iron and phosphorus rather than sunlight. The authors suggest that the tsunami triggered by impact would have stirred up soluble iron-2 from the deep ocean and washed in phosphorus from the exposed land surface, perhaps some having been delivered by the asteroid itself. No doubt such a huge impact would have veiled the Palaeoarchaean Earth with dust that reduced sunlight for years: inimical for photosynthesising bacteria but unlikely to pose a threat to chemo-autotrophs. An unusual feature of the S2 spherule bed is that it is capped by a layer of altered crystals whose shapes suggest they were originally sodium bicarbonate and calcium carbonate. They may represent flash-evaporation of up to tens of metres of ocean water as a result of the impact. Carbonates are less soluble than salt and more likely to crystallise during rapid evaporation of the ocean surface than would NaCl.
Time line of possible events following a huge asteroid impact during the Palaeoarchaean. (Credit: Drabon, N. et al. Fig 8)
So it appears that early extraterrestrial bombardment in the early Archaean had the opposite effect to the Chicxulub impactor that devastated the highly evolved life of the late Mesozoic. Many repeats of such chaos during the Palaeoarchaean could well have given a major boost to some forms of early, chemo-autotrophic life, while destroying or setting back evolutionary attempts at photo-autotrophy.
The roof lifted for palaeoanthropologists in October 2004 when news emerged of a fossil from Liang Bua cave on Flores in the Indonesian archipelago. It was an adult female human skull about a third the size of those of anatomically modern humans (see: The little people of Flores, Indonesia; October 2004). Immediately it was dubbed ‘Hobbit’, and from the start controversy raged around this diminutive human. The cave layer contained evidence of fire and sophisticated tools as well as bones of giant rats and minute elephants, presumed to be staple prey for these little people. Despite having brains about the size of a grapefruit – as did australopithecines – the little people challenged our assumptions about intelligence. Preliminary dating from 95 to 17 ka suggested they may have cohabited Indonesia with both H. erectus and AMH. Indeed, modern people of Flores tell legends of the little people they call Ebo Go Go. Like both their ancestors must have crossed treacherous straits between the Indonesian islands, which existed even when global sea level was drawn down by polar icecaps. Once an early suggestion that the original find was the skull of a deformed, microcephalic individual had been refuted by further finds in Flores, scientists turned to natural selection of small stature through living on a small island with limited resources – similar to the tiny elephant Stegodon and other island faunas elsewhere. By 2007, it had become clear from other, similar fossils that they were definitely a distinct species Homo floresiensis (see: Now we can celebrate the ‘Hobbits’! November 2007) with several anatomical similarities to H. erectus. Then more sophisticated dating revealed that the Flores cave sediments containing their fossils and tools spanned 100 to 60 ka, well before AMH reached Indonesia. By 2018 their arrival on Flores, marked by a mandible fragment and 6 teeth in sediments from sediment excavation at Mata Menge 70 km east of Luing Bua, had been pushed back to 773 ka. At the new site stone tools were found in even earlier sediments (1.02 Ma). In 2019 evidence emerged that isolated island evolution in the Philippines had produced similar small descendants (H. luzonensis) by around 67 ka.
Artist’s impression of Homo floresiensis with giant rat. (Credit: Box of Oddities podcast)
The latest development is the finding of a fragment of an adult humerus (an arm bone) in the Mata Menge excavations that had yielded the oldest dates for Homo floresiensis fossils (Kaifu, Y. and 12 others 2024. Early evolution of small body size in Homo floresiensis. Nature Communications, v. 15, article number 6381; DOI: 10.1038/s41467-024-50649-7). Comparing the teeth and arm-bone fragment with an intact adult from Liang Bua suggests that the earliest known ancestors of Homo floresiensis were even smaller. The teeth, albeit much smaller, resemble those of Indonesian specimens of H. erectus. That observation helps to rule out earlier speculation that the tiny people of Flores descended from the earliest humans from Africa (H. habilis) that were about the same size, but more than twice as old (2.3 to 1.7 Ma). The evidence points more plausibly towards their evolution from Asian H. erectus, who arrived in Java around 1.1 million years ago. Having solved the issue of ‘island hopping’ to reach Java a group of Asian H. erectus could have found their way to Flores. That island’s biological resources may not have met the survival requirements of a much larger human ancestor but evolution in isolation kept the arrivals alive. Within 300 ka, and perhaps much less for a small population, survival of smaller offspring allowed them a very long and apparently quite comfortable stay on the island. Though diminished in stature, they demonstrated the survival strategies conferred by being smart.
In 1961 ten scientists interested in a search for extra-terrestrial intelligence met at Green Bank, West Virginia, USA, none of whom were geologists or palaeontologists. The participants called themselves “The Order of the Dolphin”, inspired by the thorny challenge of discovering how small cetaceans communicated: still something of a mystery. To set the ball rolling, Frank Drake an American astrophysicist and astrobiologist, proposed an algorithm aimed at forecasting the number of planets elsewhere in our galaxy on which ‘active, communicative civilisations’ (ACCs) might live. The Drake Equation is formulated as:
ACCs = R* · fp · ne · fl · fi · fc · L
where R* = number of new stars formed per year, fp = the fraction of stars with planetary systems, ne = the average number of planets that could support life (habitable planets) per planetary system, fl = the fraction of habitable planets that develop primitive life, fi = the fraction of planets with life that evolve intelligent life and civilizations, fc = the fraction of civilizations that become ACCs,L = the length of time that ACCs broadcast radio into space. A team of then renowned scientists from several disciplines discussed what numbers to attach to these parameters. Their ‘educated guesses’ were: R* – one star per year; fp – one fifth to one half of all stars will have planets; ne – 1 to 5 planets per planetary system will be habitable; of which 100% will develop life (fl) and 100% (fi) will eventually develop intelligent life and civilisations; of those civilisations 10 to 20 % (fc) will eventually develop radio communications; which will survive for between a thousand years and 100 Ma (L). Acknowledging the great uncertainties in all the parameters, Drake inferred that between 103 and 108 ACCs exist today in the Milky Way, which is ~100 light years across and contains 1 to 4 x 1011 stars).
Today the values attached to the parameters and the outcomes seem absurdly optimistic to most people, simply because, despite 4 decades of searching by SETI there have been no signs of intelligible radio broadcasts from anywhere other than Earth and space probes launched from here. This is humorously referred to as the Fermi Paradox. There are however many scientists who still believe that we are not alone in the galaxy, and several have suggested reasons why nothing has yet been heard from ACCs. Robert Stern of the University of Texas (Dallas), USA and Taras Gerya of ETH-Zurich, Switzerland have sought clues from the history of life on Earth and that of the inorganic systems from which it arose and in which it has evolved that bear on the lack of any corrigible signals in the 63 years since the Drake Equation (Stern, R.J & Gerya, T.V. 2024. The importance of continents, oceans and plate tectonics for the evolution of complex life: implications for finding extraterrestrial civilizations. Nature (Scientific Reports), v. 14, article 8552; DOI: 10.1038/s41598-024-54700-x – definitely worth reading). Of course, Stern and Gerya too are fascinated by the scientific question as to whether or not there are ‘active, communicative civilisations’ elsewhere in the cosmos. Their starting point is that the Drake Equation is either missing some salient parameters, or that those it includes are assigned grossly optimistic magnitudes.
Life seems to have been present on Earth 3.8 Ga ago but multicelled animals probably arose only in the Late Neoproterozoic since 1.0 Ga ago. So here it has taken a billion years for their evolution to achieve terrestrial ACC-hood. Stern and Gerya address what processes favour life and its rapid evolution. Primarily, life depends on abundant liquid water: i.e. on a planet within the ‘Goldilocks Zone’ around a star. The authors assume a high supply of bioactive compounds – organic carbon, ammonium, ferrous iron and phosphate to watery environments. Phosphorus is critical to their scenario building. It is most readily supplied by weathering of exposed continental crust, but demands continual exposure of fresh rock by erosion and river transport to maintain a steady supply to the oceans. Along with favourable climatic conditions, that can only be achieved by an oxidising environment that followed the Great Oxidation Event (2.4 to 2.1 Ga) and continual topographic rejuvenation by plate tectonics.
A variety of Earth-logs posts have discussed various kinds of evidence for the likely onset of plate tectonics, largely focussing on the Hadean and Archaean. Stern and Gerya prefer the Proterozoic Eon that preserves more strands of relevant evidence, from which sea-floor spreading, subduction and repeated collision orogenies can confidently be inferred. All three occur overwhelmingly in Neoproterozoic and Phanerozoic times. Geologists often refer to the whole of the Mesoproterozoic and back to about 2.0 Ga in the Palaeoproterozoic as the ‘Boring Billion’ during which carbon isotope data suggest very little change in the status of living processes: they were present but nothing dramatic happened after the Great Oxidation Event. ‘Hard-rock’ geology also reveals far less passive extensional events that indicate continental break-up and drift than occur after 1.0 Ga and to the present. It also includes a unique form of magmatism that formed rocks dominated by sodium-rich feldspar (anorthosites) and granites that crystallised from water-poor magmas. They are thought to represent build-ups of heat in the mantle unrelieved by plate-tectonic circulation. Before the ‘Boring Billion’ such evidence as there is does point to some kind of plate motions, if not in the modern style.
How different styles of tectonics influence living processes differently: a single stagnant ‘lid’ versus plate tectonics. (Credit: Stern and Gerya, Fig 2)
Stern and Gerya conclude that the ‘Boring Billion’ was dominated by relative stagnation in the form of lid tectonics. They compare the influence of stagnant ‘lid’ tectonics on life and evolution with that of plate tectonics in terms of: bioactive element supply; oxygenation; climate control; habitat formation; environmental pressure (see figure). In each case single lid tectonics is likely to retard life and evolution, whereas plate tectonics stimulates them as it has done from the time of Snowball Earth and throughout the Phanerozoic. Only one out of 8 planets that orbit the sun displays plate tectonics and has both oceans and continents. Could habitable planets be a great deal rarer than Drake and his pals assumed? [look at exoplanets in Wikipedia] Whatever, Stern and Gerya suggest that the seemingly thwarted enthusiasm surrounding the Drake Equation needs to be tempered by the addition of two new terms: the fraction of habitable exoplanets with significant continents and oceans (foc)and the fraction of them that have experienced plate tectonics for at least half a billion years (fpt). They estimate foc to be on the order of 0.0002 to 0.01, and suggest a value for fpt of less than 0.17. Multiplied together yields value between less than 0.00003 and 0.002. Their incorporation in the Drake Equation drastically reduces the potential number of ACCs to between <0.006 and <100,000, i.e. to effectively none in the Milky Way galaxy rising to a still substantial number
There are several other reasons to reject such ‘ball-parking’ cum ‘back-of-the-envelope’ musings. For me the killer is that biological evolution can never be predicted in advance. What happened on our home world is that the origin and evolution of life have been bound up with the unique inorganic evolution of the Solar System and the Earth itself over more than 4.5 billion years. That ranges in magnitude from the early collision with another, Mars-sized world that reset the proto-Earth’s geochemistry and created a large moon whose gravity has cycled the oceans through tides and changed the length of the day continually for almost the whole of geological history. At least once, at the end of the Cretaceous Period, a moderately sized asteroid in unstable orbit almost wiped out life at an advanced stage in its evolution. During the last quarter billion years internally generated geological forcing mechanisms have repeatedly and seriously stressed the biosphere in roughly 36 Ma cycles (Boulila, S. et al. 2023. Earth’s interior dynamics drive marine fossil diversity cycles of tens of millions of years. Proceedings of the National Academy of Sciences, v. 120 article e2221149120; DOI: 10.1073/pnas.2221149120). Two outcomes were near catastrophic mass extinctions, at the ends of the Permian and Triassic Periods, from which life struggled to continue. As well as extinctions, such ‘own goals’ reset global ecosystems repeatedly to trigger evolutionary diversification based on the body plans of surviving organisms.
Such unique events have been going on for four billion years, including whatever triggered the Snowball Earth episodes that accompanied the Great Oxygenation Event around 2.4 Ga and returned to coincide with the rise of multicelled animals during the Cryogenian and Ediacaran Periods of the Late Neoproterozoic. For most of the Phanerozoic a background fibrillation of gravitational fields in the Solar System has occasionally resulted in profound cycling between climatic extremes and their attendant stresses on ecosystems and their occupants. The last of these coincided with the evolution of humanity: the only creator of an active, communicative civilisation of which we know anything. But it took four billion years of a host of diverse vagaries, both physical and biological to make such a highly unlikely event possible. That known history puts the Drake Equation firmly in its place as the creature of a bunch of self-publicising and regarding, ambitious academics who in 1961 basically knew ‘sweet FA’. I could go on … but the wealth of information in Stern and Gerya’s work is surely fodder for a more pessimistic view of other civilisations in the cosmos.
Someone – I forget who – provided another, very practical reason underlying the lack of messages from afar. It is not a good idea to become known to all and sundry in the galaxy, for fear that others might come to exploit, enslave and/or harvest. Earth is still in a kind of imperialist phase from which lessons could be drawn!
You can follow my ‘reportage’ on the long running story of the Snowball Earth events during the Neoproterozoic Cryogenian Period (850 to 635 Ma) since 2000 through the index to annual Palaeoclimatology logs (15 posts). Once these dramatic events were over sedimentary rocks deposited around the world during the Ediacaran Period (635 to 541 Ma) record the sudden appearance of large-bodied fossils: the first multicellular animals. This explosion from slimy biofilms and colonies of single-celled prokaryotes and eukaryotes laid the basis for the myriad ecological niches that have characterised Planet Earth ever since. The change saw specialised eukaryote cells (see: The rise of the eukaryotes; December 2017), whose precursors had originated in single-celled forms, begin to cooperate inthe development of complex tissues, organs, and organ systems to form bodies rather than just cell walls. The pulsating evolution, diversification and repeated extinction that followed during the last one tenth of geological time shaped a planet that is unique in the Solar System and possibly in the galaxy, if not the entire universe. The simple biosphere that preceded it, on the other hand, may have emerged on innumerable rocky planets blessed with liquid water to survive little changed for billions of years, as have Earths’ prokaryotes, the Archaea and Bacteria.
Artist’s impression of the Ediacaran Fauna (credit: Science)
The Ediacaran biological revolution followed repeated changes in the geochemistry of the oceans, which carbon isotope data from the Cryogenian and Ediacaran suggest to have ‘gone haywire’. This turmoil involved dramatic changes in the cycling of sulfur and phosphorus that help ‘fertilise’ the marine food chain and in the production of oxygen by photosynthesis that is essential for metazoan animals. The episodes when the Earth was iced over reduced the availability of nutrients through decreased rates of ocean-floor burial of dead organisms. Such Snowball events would also have reduced penetration of sunlight in the oceans. Less photosynthesis would not only have reduced oxygen production but also the amounts of autotrophic organisms. Furthermore, decreased water temperature would have increased its viscosity thereby slowing the spread of nutrients. The food chain for heterotrophs was decimated. Each Snowball event ended with warming, ice-free conditions so that the marine biosphere could burgeon
A great deal of data and numerous theories have accumulated since the Snowball concept was first mooted, but there has been little progress in understanding the rise of multi-celled life. Four geoscientists from the Massachusetts Institute of Technology, the Santa Fe Institute and the University of Colorado (Boulder), USA have developed an interesting hypothesis for how this enormous evolutionary step may have developed (Crockett, W.W. et al. 2024. Physical constraints during Snowball Earth drive the evolution of multicellularity. Proceedings of the Royal Society B: Biological Sciences, v. 291; DOI: 10.1098/rspb.2023.2767). The concatenation of huge events during the Cryogenian and Ediacaran presented continually changing patterns of selective pressures on simple organisms that preceded that time period. Crockett et al. review them in the light of fundamental biology to suggest how multicellular animals emerged as the Ediacara Fauna. Intuitively, such harsh conditions suggest at worst mass, even complete, extinction, at best a general reduction in size of all organism to cope with scarce resources. That the size of eukaryotes should have grown hugely goes against the grain of most biologists’ outlook.
The authors consider the crucial factor to be fundamental differences between prokaryotes and early eukaryotes. Prokaryote cells are very small, and whether autotrophs of heterotrophs they absorb nutrients through their walls by diffusion. Single-celled eukaryotes are far larger than prokaryotes and typically have a flagellum or ‘tail’ so that they can move independently and more easily gather resources. Crockett et al. used computer modelling to simulate the type of life form that could grow and thrive under Snowball conditions. They found that prokaryotes could only grow smaller, being ‘stunted’ by scarce resources. On the other hand eukaryotes would be better equipped to gather resources, the more so if they adopted a simple multicellular form – a hollow, self-propelled sphere about the size of a pea, which the authors dub a choanoblastula. Although no such form is known today, it does resemble the green Volvox algae, and plausibly could have evolved further to the simple forms of the Ediacaran fauna. The next task is either to find a fossil of such an organism, or to grow one.
Charles Darwin famously suggested that humans evolved from apes, and since great apes (chimpanzees, bonobos and gorillas) live in Africa he reckoned it was probably there that the human ‘line’ began. Indeed, the mitochondrial DNA of chimpanzees (Pan troglodytes) is the closest to that of living humans. Palaeoanthropology in Africa has established evolutionary steps during the Pleistocene (2.0 to 0.3 Ma) by early members of the genus Homo: H. habilis, H. ergaster, H. erectus; H. heidelbergensis and the earliest H. sapiens. Members of the last three migrated to Eurasia, beginning around 1.8 Ma with the individuals found at Dmanisi in Georgia. The earliest African hominins emerged through the Late Miocene (7.0 to 5.3 Ma): Sahelanthropus tchadensi, Orrorin tugenensis and Ardipthecus kadabba. Through the Pliocene (5.3 to 2.9 Ma) and earliest Pleistocene two very distinct hominin groups appeared: the ‘gracile’ australopithecines (Ardipithecus ramidus; Australopithecus anamensis; Au. afarensis; Au. africanus; Au. sediba) and the ‘robust’ paranthropoids (Paranthropus aethiopicus; P. robustus and P. boisei). The last of the paranthropoids cohabited East Africa with early homo species until around 1.4 Ma. Most of these species have been covered in Earth-logs and an excellent time line of most hominin and early human fossils is hosted by Wikipedia.
All apes, including ourselves, and fossil examples are members of the Family Hominidae (hominids) which refers to the entire world. A Subfamily (Homininae) refers to African apes, with two Tribes. One, the Gorillini, refers to the two living species of gorilla. The other is the Hominini (hominins) that includes chimpanzees, living humans and all fossils believed to be on the evolutionary line to Homo. The Tribe Hominini is defined to have descended from the common ancestor of modern humans and chimps, and evolved only in Africa. As the definition of hominins stands, it excludes other possibilities! The Miocene of Africa before 7.2 Ma ‘goes cold’ as regards the evolution of hominins. There are, however fossils of other African apes in earlier Miocene strata (8 to 18 Ma) that have been assigned to the Family Hominidae, i.e. hominids, of which more later.
Much has been made of using a ‘molecular clock’ to hint at the length of time since the mtDNA of living humans and chimps began to diverge from their last common ancestor. That is a crude measure at it depends entirely on assuming a fixed rate at which genetic mutation in primates take place. Many factors render it highly uncertain, until ancient DNA is recovered from times before about 400 ka, if ever. The approach suggests a range from 7 to 10 Ma, yet the evolutionary history of chimps based on fossils is practically invisible: the earliest fossil of a member of genus Pan is from the Middle Pleistocene (1.2 to 0.8 Ma) of Kenya. Indeed, we have little if any clue about what such a common ancestor looked like or did. So the course of human evolution relies entirely on the fossil sequence of earlier African hominins and comparing their physical appearances. Each species in the African time line displays two distinctive features. All were bipedal and had small canine teeth. Modern chimps habitually use knuckle walking except when having to cross waterways. As with virtually all other primates, fossil or living, male chimps have large, threatening canines. In the absence of ancient DNA from fossils older than 0.4 Ma these two features present a practical if crude way of assessing to when and where the hominin time line leads.
In 2002 a Polish geologist on holiday at the beach at Trachilos on Crete discovered a trackway on a bedding plane in shallow-marine Miocene sediments. It had been left by what seems to have been a bipedal hominin. Subsequent research was able to date the footprints to about 6.05 Ma. Though younger than Sahelanthropus, the discovery potentially challenges the exclusivity of hominins to Africa. Unsurprisingly, publication of this tentative interpretation drew negative responses from some quarters. But the discovery helped resurrect the notion that Africa may have been colonised in the Miocene by hominins that had evolved in Europe. That had been hinted at by the 1872 excavation of Oreopithecus bamboliifrom an Upper Miocene (~7.6 Ma) lignite mine in Tuscany, Italy – a year after publication of Darwin’s The Descent of Man.
Lignites in Tuscany and Sardinia have since yielded many more specimens, so the species is well documented. Oreopithecus could walk on two legs, its hands were capable of a precision grip and it had relatively small canines. Its Wikipedia entry cautiously refers to it as ‘hominid’ – i.e. lumped with all apes to comply with current taxonomic theory (above). In 2019 another fascinating find was made in a clay pit in Bavaria, Germany. Danuvius guggenmosi lived 11.6 Ma ago and fossilised remains of its leg- and arm bones suggested that it could walk on two legs: it too may have been on the hominin line. But no remains of Danuvius’s skull or teeth have been found. There is now an embarrassment of riches as regards Miocene fossil apes from Europe and the Eastern Mediterranean (Sevim-Erol, A. and 8 others 2023. A new ape from Türkiye and the radiation of late Miocene hominines. Nature Communications Biology, v. 6, article 842.; DOI: 10.1038/s42003-023-05210-5). A number of them closely resemble the earliest fossil hominins of Africa, but most predate the hominin record there by several million years.
Phylogenetic links between fossils assigned to Hominidae found in Africa and north of the Mediterranean Sea. (Credit: Sevim-Erol et al. 2023, Fig 5)
Ayla Sevim-Erol of Ankara University, Turkiye and colleagues from Turkiye, Canada and the Netherlands describe a newly identified Miocene genus, Anadoluvius, which they place in the Subfamily Homininae dated to around 8.7 Ma. Fragments of crania and partial male and female mandibles from Anatolia show that its canines were small and comparable with those of younger African hominins, such as Ardipithecus and Australopithecus. But limb bones are yet to be found. Around the size of a large male chimpanzee, Anadoluvius lived in an ecosystem remarkably like the grasslands and dry forests of modern East Africa, with early species of giraffes, wart hogs, rhinos, diverse antelopes, zebras, elephants, porcupines, hyenas and lion-like carnivores. Sevim-Erol et al. have attempted to trace back hominin evolution further than is possible with African fossils. They compare various skeletal features of different fossils and living genera to assess varying degrees of similarity between each genus, applied to 23 genera. These comprised 7 hominids from the African Miocene, 2 early African hominins (Ardipithecus and Orrorin) and 10 Miocene hominids from Europe and the Eastern Mediterranean. They also assessed similarities with 4 living genera, Homo, orang utan (Pongo), gorilla and chimp (Pan).
The resulting phylogeny shows close morphological links within a cluster (green ‘pools’ on diagram) of non-African hominids with the African hominins, gorillas, humans and chimps. There are less-close relations between that cluster and the earlier Miocene hominids of Africa (blue ‘pool’) and the possible phylogeny of orang utans (orange ‘pool’). Sevim-Erol et al. note that African hominins are clearly more similar and perhaps more closely related to the fossils of Europe and the Eastern Mediterranean than they are to Miocene African hominids. This suggests that evolution among the non-African hominids ceased around the end of the Miocene Epoch north of the Mediterranean Sea. But it may have continued in Africa. Somehow, therefore, it became possible late in Miocene times for hominids to migrate from Europe to Africa. Yet the earlier, phylogenetically isolated African hominids seem to have ‘crashed’ at roughly the same time. Such a complex scenario cannot be supported by phylogenetic studies alone: it needs some kind of ecological impetus.
The Mediterranean Basin at the end of the Miocene Epoch when the only water was in the deepest parts of the basin. (Credit: Wikipedia, Creative Commons)
Following a ‘mild’ tectonic collision between the African continent and the Iberian Peninsula during the late Miocene connection between the Atlantic Ocean and the Mediterranean Sea was blocked from 6.0 to 5.3 Ma. Except for its deepest parts, seawater in the Mediterranean evaporated away to leave thick salt deposits. Rivers, such as the Rhône, Danube, Dneiper and Nile, shed sediments into the exposed basin. For 700 ka the basin was a fertile, sub-sea level plain, connecting Europe and North Africa over and E-W distance of 3860 km. There was little to stop the faunas of Eurasia and Africa migrating and intermingling, at a critical period in the evolution of the Family Hominidae. One genus presented with the opportunity was quite possibly the last common ancestor of all the hominins and chimps. The migratory window vanished at the end of the Miocene when what became the Strait of Gibraltar opened at 5.3 to allow Atlantic water. This resulted in the stupendous Zanclean flood with a flow rate about 1,000 times that of the present-day Amazon River. An animation of these events is worth watching
The largest volcanic eruption during the 2.5 million year evolution of the genius Homo, about 74 thousand years (ka) ago, formed a huge caldera in Sumatra, now filled by Lake Toba. A series of explosions lasting just 9 to 14 days was forceful enough to blast between 2,800 to 6,000 km3 of rocky debris from the crust. An estimated 800 km3 was in the form of fine volcanic ash that blanketed South Asia to a depth of 15 cm. Thin ash layers containing shards of glass from Toba occur in marine sediments beneath the Indian Ocean, the Arabian and South China Seas. Some occur as far off as sediments on the floor of Lake Malawi in southern Africa. A ‘spike’ of sulfates is present at around 74 ka in a Greenland ice core too. Stratospheric fine dust and sulfate aerosols from Toba probably caused global cooling of up to 3.5 °C over a modelled 5 years following the eruption. To make matters worse, this severe ‘volcanic winter’ occurred during a climatic transition from warm to cold caused by changes in ocean circulation and falling atmospheric CO2 concentration, known as a Dansgaard-Oeschger event.
There had been short-lived migrations of modern humans out of Africa into the Levant since about 185 ka. However, studies of the mitochondrial DNA (mtDNA) of living humans in Eurasia and Australasia suggest that permanent migration began about 60 ka ago. Another outcome of the mtDNA analysis is that the genetic diversity of living humans is surprisingly low. This suggests that human genetic diversity may have been sharply reduced globally roughly around the time of the Toba eruption. This implies a population bottleneck with the number of humans alive at the time to the order of a few tens of thousands (see also: Toba ash and calibrating the Pleistocene record; December 2012). Could such a major genetic ‘pruning’ have happened in Africa? Over six field seasons, a large team of geoscientists and archaeologists drawn from the USA, Ethiopia, China, France and South Africa have excavated a rich Palaeolithic site in the valley of the Shinfa River, a tributary of the Blue Nile in western Ethiopia. Microscopic studies of the sediments enclosing the site yielded glass shards whose chemistry closely matches those in Toba ash, thereby providing an extremely precise date for the human occupation of the site: during the Toba eruption itself (Kappelman, Y. and 63 others 2024. Adaptive foraging behaviours in the Horn of Africa during Toba supereruption. Nature, v. 627; DOI: 10.1038/s41586-024-07208-3).
Selection of possible arrowheads from the Shinfa River site (Credit: Kappelman et al.; Blue Nile Survey Project)
The artifacts and bones of what these modern humans ate suggest a remarkable scenario for how they lived. Stone tools are finely worked from local basalt lava, quartz and flint-like chalcedony found in cavities in lava flows. Many of them are small, sharp triangular points, some of which show features consistent with their use as projectile tips that fractured on impact; they may be arrowheads, indeed the earliest known. Bones found at the site are key pointers to their diet. They are from a wide variety of animal, roughly similar to those living in the area at present: from monkeys to giraffe, guinea fowl to ostrich, and even frogs. There are remains of many fish and freshwater molluscs. Although there are no traces of plant foods, clearly those people who loved through the distant effects of Toba were well fed. Although a period of global cooling may have increased aridity at tropical latitudes in Africa, the campers were able to devise efficient strategies to obtain victuals. During wet seasons they lived off terrestrial prey animals, and during the driest times ate fish from pools in the river valley. These are hardly conditions likely to devastate their numbers, and the people seem to have been technologically flexible. Similar observations were made at the Pinnacle Point site in far-off South Africa in 2018, where Toba ash is also present. Both sites refute any retardation of human cultural progress 74 ka ago. Rather the opposite: people may have been spurred to innovation, and the new strategies may have allowed them to migrate more efficiently, perhaps along seasonal drainages. In this case that would have led them or their descendants to the Nile and a direct route to Eurasia; along ‘blue highway’ corridors as Kappelman et al. suggest.
Yet the population bottleneck implied by mtDNA analyses is only vaguely dated: it may have been well before or well after Toba. Moreover, there is a 10 ka gap between Toba and the earliest accurately dated migrants who left Africa – the first Australians at about 65 ka. However, note that there is inconclusive evidence that modern humans may have occupied Sumatra by the time of the eruption. Much closer to the site of the eruption in southeast India, stone artifacts have been found below and above the 74 ka datum marked by the thick Toba Ash. Whether these were discarded by anatomically modern humans or earlier migrants such as Homo erectus remains unresolved. Either way, at that site there is no evidence for any mass die-off, even though conditions must have been pretty dreadful while the ash fell. But that probably only lasted for little more than a month. If the migrants did suffer very high losses to decrease the genetic diversity of the survivors, it seems just as likely to have been due to attrition on an extremely lengthy trek, with little likelihood of tangible evidence surviving. Alternatively, the out-of-Africa migrants may have been small in number and not fully representative of the genetic richness of the Africans who stayed put: a few tens of thousand migrants may not have been very diverse from the outset.
Championed as the earliest commonly found human species and, apart from anatomically modern humans (AMH), the most widespread through Africa and Eurasia. It also endured longer (~1.75 Ma) than any other hominin species, appearing first in East Africa around 2 Ma ago, the youngest widely accepted fossil – found in China – being around 250 ka old. The ‘erects’ arguably cooked their food and discovered the use of fire 1.7 to 2 Ma ago. The first fossils discovered in Java by Eugene Dubois are now known to be associated with the oldest-known art (430 to 540 ka) The biggest issue surrounding H. erectus has been its great diversity, succinctly indicated by a braincase capacity ranging from 550 to 1250 cm3: from slightly greater than the best endowed living apes to within the range of AMH. Even the shape of their skulls defies the constraints placed on those of other hominin species. For instance, some have sagittal crests to anchor powerful jaw muscles, whereas others do not. What they all have in common are jutting brow ridges and the absence of chins along with all more recently evolved human species, except for AMH.
This diversity is summed up in 9 subspecies having been attributed to H. erectus, the majority by Chinese palaeoanthropologists. Chinese fossils from over a dozen sites account for most of the anatomical variability, which perhaps even includes Denisovans, though their existence stems only through the DNA extracted from a few tiny bone fragments. So far none of the many ‘erect’ bones from China have been submitted to genetic analysis, so that connection remains to be tested. Several finds of diminutive humans from the Indonesian and Philippine archipelagos have been suggested to have evolved from H. erectus in isolation. All in all, the differences among the remains of H. erectus are greater than those used to separate later human species, i.e. archaic AMH, Neanderthals, Denisovans, H. antecessor etc. So it seems strange that H. erectus has not been split into several species instead of being lumped together, in the manner of the recently proposed Homo bodoensis. Another fossil cranium has turned up in central China’s Hubei province, to great excitement even though it has not yet been fully excavated (Lewis, D. 2022. Ancient skull uncovered in China could be million-year-old Homo erectus. Nature News 29 November 2022; DOI: 10.1038/d41586-022-04142-00; see also a video). Chances are that it too will be different from other examples. It also presents a good excuse to consider H. erectus.
Cranium of a Chinese Homo erectus, somewhat distorted by burial, from a site close to the latest find. (Credit: Hubei Museum, Wuhan, China)
The complications began in Africa with H. ergaster, the originator of the bifacial or Acheulean multi-purpose stone tool at around 1.6 Ma (see: Flirting with hand axes; May 2009), the inventor of cooking and discoverer of the controlled use of fire. ‘Action Men’ were obviously smarter than any preceding hominin, possibly because of an increase of cooked protein and plant resources that are more easily digested than in the raw state and so more available for brain growth. The dispute over nomenclature arose from a close cranial similarity of H. ergaster to the H. erectus discovered in Java in the 19th century: H. erectus ergaster is now its widely accepted name. In 1991-5 the earliest recorded hominins outside Africa were found at Dmanisi, Georgia, in sediments dated at around 1.8 Ma (see: First out of Africa; November 2003) Among a large number of bones were five well-preserved skulls, with brain volumes less than 800 cm3 (see: An iconic early human skull; October 2013). These earliest known migrants from Africa were first thought to resemble the oldest humans (H.habilis) because of their short stature, but now are classified as H. erectus georgicus. They encapsulate the issue of anatomical variability among supposed H. erectus fossils, each being very different in appearance, one even showing ape-like features. Another had lost all teeth from the left side of the face, yet had survived long after their loss, presumably because others had cared for the individual.
The great variety of cranial forms of the Asian specimens of H. erectus may reflect a number of factors. The simplest is that continuous presence of a population there for as long as 1.5 Ma inevitably would have resulted in at least as much evolution as stemmed from the erects left behind in Africa, up to and including the emergence of AMH in North Africa about 300 ka ago. If contact with the African human population was lost after 1.8 Ma, the course of human evolution in Africa and Asia would clearly have been different. But that leaves out the possibility of several waves of migrants into Asia that carried novel physiological traits evolved in Africa to mix with those of earlier Asian populations. From about 1 Ma ago a succession of migrations from Africa populated Europe – H. antecessor, H. heidelbergensis, and Neanderthals and then AMH. So a similar succession of migrants could just as well have gone east instead of west on leaving Africa. Asia is so vast that migration may have led different groups to widely separated locations, partially cut-off by mountain ranges and deserts so that it became very difficult for them to maintain genetic contact. Geographic isolation of small groups could lead to accelerated evolution, similar to that which may have led to the tiny H. floresiensisandH. luzonensisdiscovered on Indonesian and Philippine islands.
Another aspect of the Asian continent is its unsurpassed range of altitude, latitude and climate zones. Its ecologically diversity offers a multitude of food resources, and both climate and elevation differences pose a range of potential stresses to which humans would have had to adapt. The major climate cycles of the Pleistocene would have driven migration across latitudes within the continent, thereby mixing groups with different physical tolerances and diets to which they had adapted. Equally, westward migration was possible using the Indo-Gangetic plains and the shore of the Arabian Sea: yet more opportunities for mixing between established Asians and newly arrived African emigrants.
The earliest known human fossils outside of Africa were found at a site near Dmanisi in Georgia, between 1991 and 2005, following the discovery there in 1984 of primitive stone tools together with early Pleistocene animal bones. The Dmanisi finds occur with those of sabre-toothed cats and giant cheetahs, and so are probably not interments or in some kind of dwelling but were probably dragged into an underground carnivore den.
The five Dmanisi skulls of Homo erectus georgicus (credits; M.S. Ponce de Leon & P.E. Zollkofer, University of Zurich)
Initially the remains were assigned to a new species – Homogeorgicus – but are now believed to be a subspecies of H. erectus. The finds are anatomically rich, with fossils of at least 5 individuals, both male and female, including 5 well-preserved skulls. Analysing them has been a long process. Details of the best preserved, indeed the most complete early Homo skull ever found, have taken 8 years since its discovery in 2005 to reach publication (Lordkipanidze, D. et al. 2013. A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, v. 342, p. 326-331, DOI: 10.1126/science.1238484).
To the surprise of palaeoanthropologists, this specimen of Homoerectus georgicus has some ape-like features, including a protruding upper jaw in a relatively large face that most resembles the oldest African H. habilis, from Ethiopia, dated at 2.3 Ma. With a braincase of 546 cm3, the skull is on the small side of H. habilis and in the range of late australopithecines. Yet, like the much younger Homo floresiensis – dubbed ‘the Hobbit’ – the association with tools, of the most basic Oldowan type, places it a cut above non-human hominins. The rest of the skeletal fossils show individuals with modern human proportions, albeit somewhat diminutive.
Surprises multiplied when comparative studies of all 5 skulls were complete. They are so different that, if found in widely separated specimens, would be placed in different species by most anatomists. Ruling out the chance association of several human species far from their Africa origins – few would suggest that up to 5 species left Africa at the same time and stuck together – a suggested explanation is that they represent a population of a human lineage in the process of evolving to a new species. The strength of this hypothesis contradicts the other recent view that several human species may have cohabited environments at different times. It also seems to throw into question the adoption of the name H. erectus for later human populations in both Africa and Eurasia: unless, as the authors tentatively suggest, there was genetic continuity and connectivity over large distances between both evolving populations

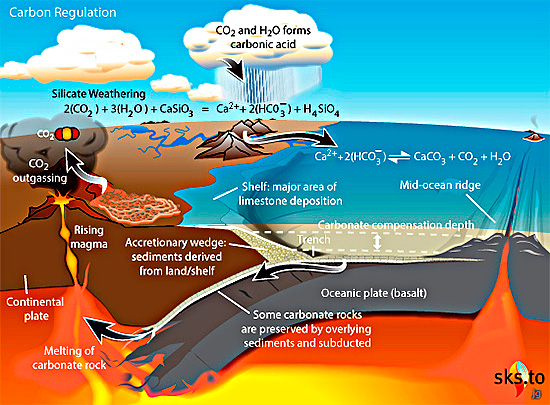


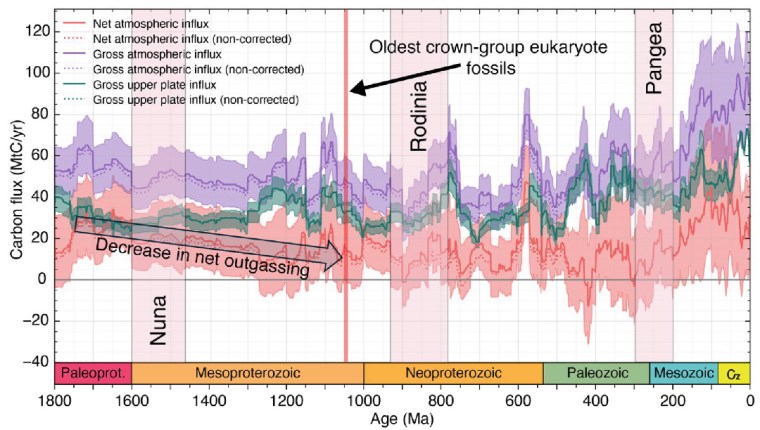
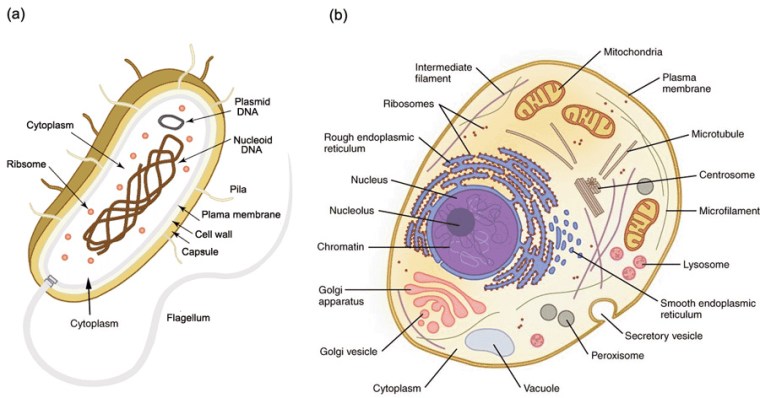











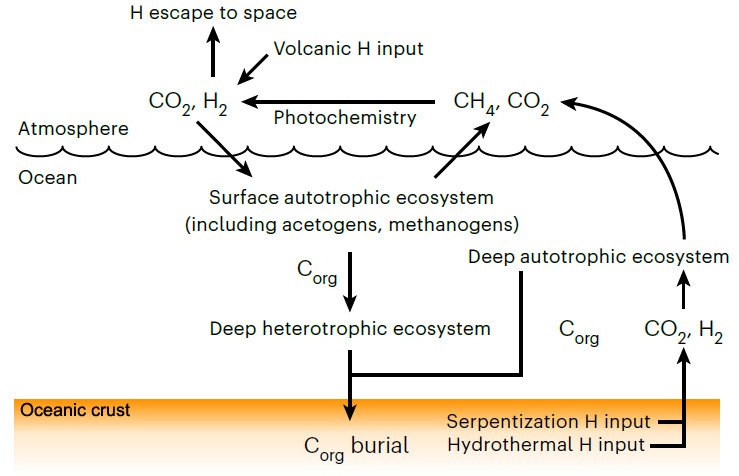



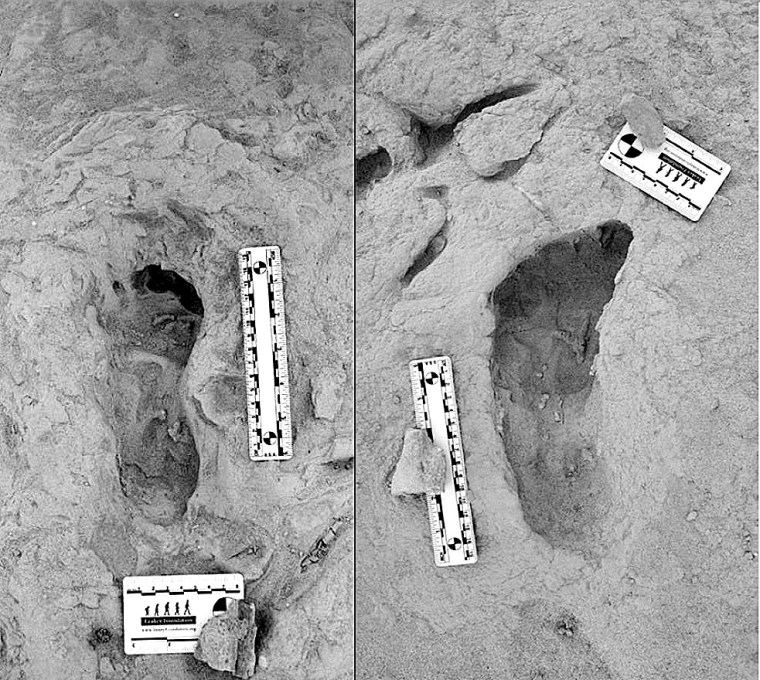



















{kind=link}