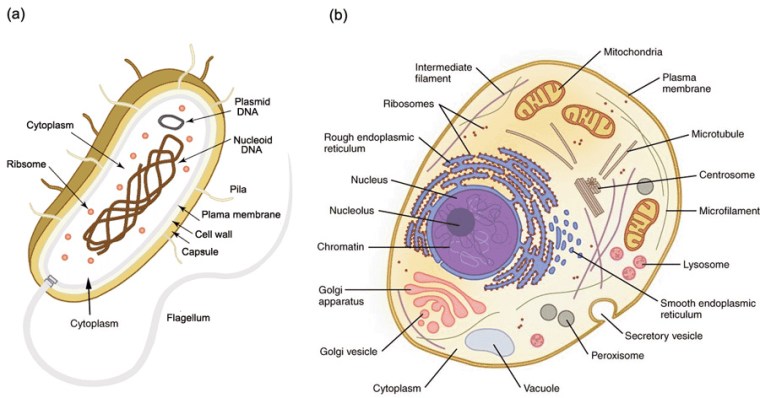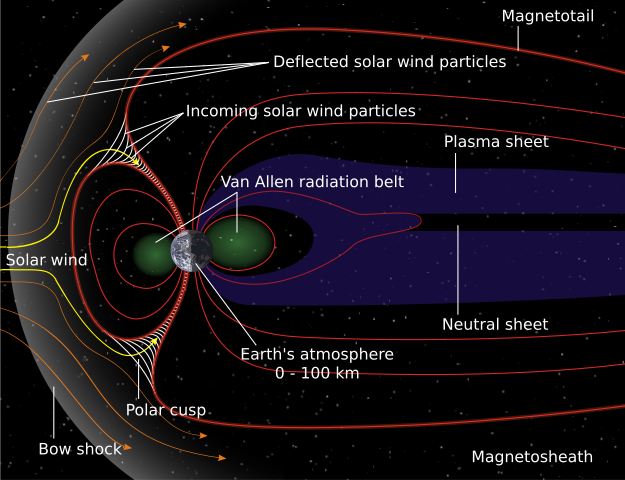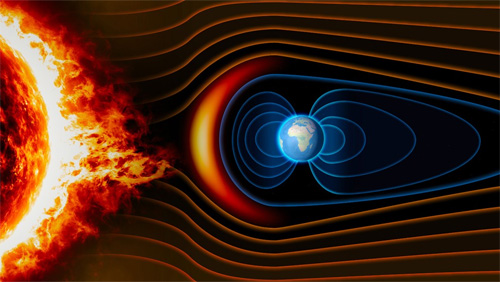
The aspect of geomagnetism that is most familiar to geologists is the magnetic field’s repeated reversals of polarity. It manifests as the magnetic ‘striping’ of modern ocean floors: key to the discovery of plate tectonics. Relatively easy to measure in intricate detail using modern magnetometers, the ‘stripes’ record plate motions back to about the Middle Jurassic (~165 Ma). There are some examples of polarity reversals from Meso- and Neoproterozoic continental rocks but not a continuum. Measuring the direction of remanent magnetism in rock samples in ancient blocks of crust allow geologists to assess their former positions and orientation relative to magnetic poles of the past, and thus evidence of ancient continental drift. Measuring the intensity of past geomagnetism – its field strength – is more difficult than either. That is critically important for Earth’s biosphere because the magnetic field deflects high-energy charged particles from the Sun and cosmic rays that would otherwise reach the surface. Without a geomagnetic field it is possible that living cells could not survive. Even with the modern protective efficiency of geomagnetism, such particles are a likely source of genetic mutations in DNA: a key requirement of biological evolution.
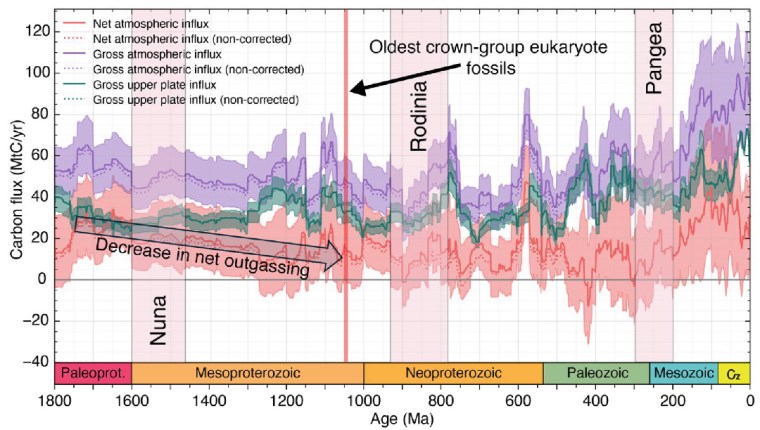
In 2022 an extremely low magnetic field strength – about 15 times less than it is today – was found in a 565 Ma-old anorthosite intrusion from Quebec, Canada that crystallised in the late Ediacaran Period. Samples from as far back as 3500 Ma in the Archaean showed a slow decline in field strength that culminated in its near vanishing in the Ediacaran. This fostered speculation that maybe increased genetic mutation by such an extraterrestrial bombardment of primitive eukaryote cells may have triggered the evolution of the multicelled Ediacara Fauna and perhaps even the first single-celled eukaryotes. A follow up in 2024 by a multinational group of scientists analysed minerals in igneous rocks from South Africa (2054 Ma) and Brazil (591 Ma) to check the magnetic-field strength in the Palaeoproterozoic Era and the Ediacaran Period (Wentao Huang and 16 others, 2024. Near-collapse of the geomagnetic field may have contributed to atmospheric oxygenation and animal radiation in the Ediacaran Period. Nature Communications: Earth and Environment, v. 5, article 207; DOI: 10.1038/s43247-024-01360-4). A more recent study by British and Brazilian geophysicists (Lloyd, S.J. et al. 2026. Earth’s magnetic field remained weak for 40 million years after the Cambrian radiation, Science Advances, v. 12, article eaeg2325; DOI: 10.1126/sciadv.aeg2325) suggested that field strength remained very low until about 505 Ma in the late Cambrian.
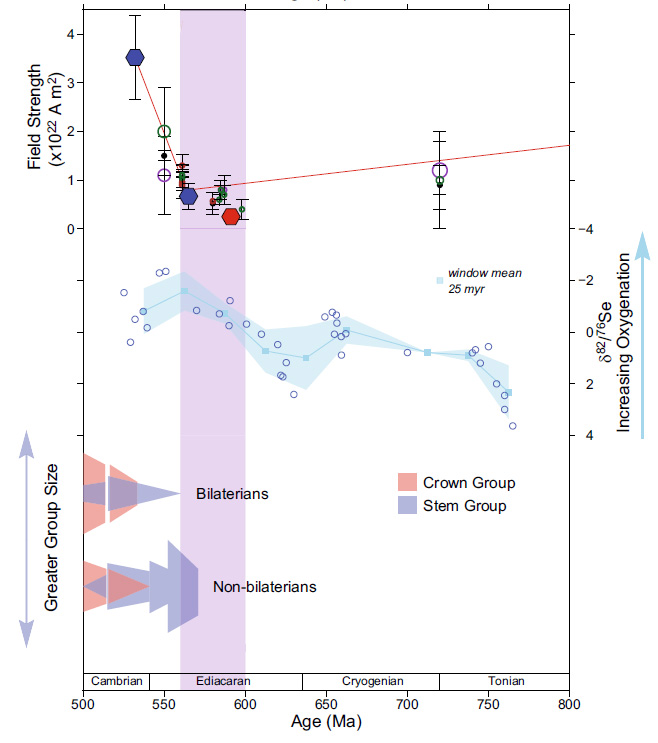
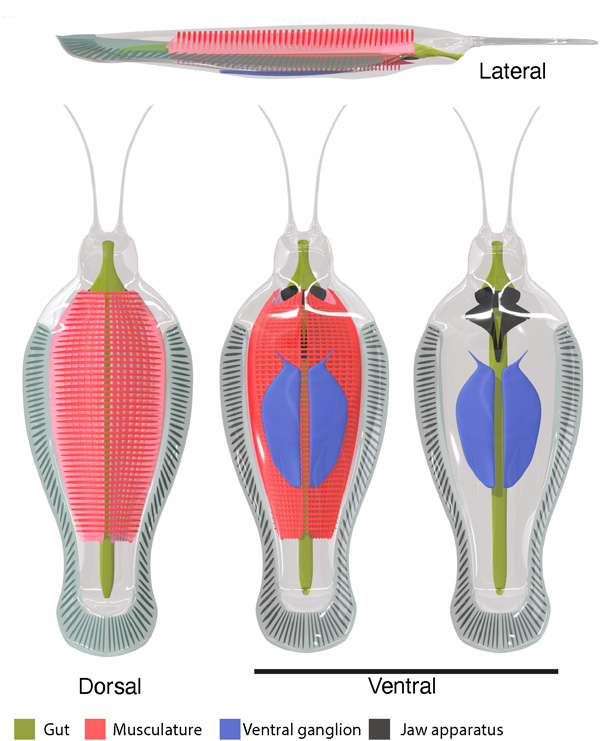
The slow decline in magnetic field strength during the Precambrian probably reflects slowing of turbulent motion in the early, completely molten core as it cooled. Eventually, at the time of very low magnetic field strength around 565 Ma the solid inner core slowly began to form through crystallisation of Fe-Ni alloy, reaching about half its present radius by the end of the Ordovician Period (~450 Ma). This roughly 100 Ma period encompasses the emergence of the Ediacara Fauna and the Cambrian ‘Explosion’ when the basal groups of almost all ‘modern’ eukaryote phyla evolved. Such a geologically swift burst of evolution now seems to be connected to more rapid genetic mutation when the Earth almost lost its ability to be shielded from radiation by its magnetic field. Natural selection had a wealth of options to test, promote or snuff out. To plagiarise the Duke of Wellington, it may have been ‘the nearest run thing you ever saw in your life’. But according to Wentao Huang et al. another beneficial factor for eukaryote evolution may have been that solar-wind bombardment could have split water into its component oxygen and hydrogen. Being so light, hydrogen would have been driven into outer space leaving Earth’s atmosphere with more oxygen, which is vital for eukaryotes.