
Hominin fossils referred to as Homo erectus have been found in Africa, Central Europe and China. Those from Africa have also been attributed to H. ergaster and by some to ‘African H. erectus’– a point of lingering dispute and confusion. The African lineage spanned a long time period after appearing around 1.7 Ma ago, possibly to as late as 0.6 Ma. The confusion deepened with the discovery of similar, well-preserved remains at Dmanisi in Georgia that are actually older (1.77–1.85 Ma) than the African specimens. But they are so anatomically diverse that the five skulls might easily be assigned to five different hominin species had they been found at separate locations. Asia is again very odd in an H. erectus context. The species was first proposed in 1891 by Eugene Dubois from remains in sediments oof the Solo River near Trinil in Java – he originally suggested the name Pithecanthropus erectus. Remarkably, the Solo sediments were dated at 53 to 27 ka in 2019, so did Homo erectus co-exist with anatomically modern humans (AMH) on Java? Similarly heavy-browed crania emerged from several sites in China. Curiously, the first complete Denisovan cranium, found near Harbin in China matched H. erectus in the eyebrow department. However, mtDNA from its dental plaque and bone proteomics tie in with those found in fragments from the eponymous Denisova Cave in Siberia and with a fragmentary mandible from Tibet. Without such evidence, and were itnot so young (146 ka), most palaeoanthropologists would probably have called the Harbin individual H. erectus, There are quite a few records of older Chinese hominin crania, dubbed H. erectus on anatomical grounds, including one dated at 1.7 Ma. There are a great many oddities and contradictions that need resolving.
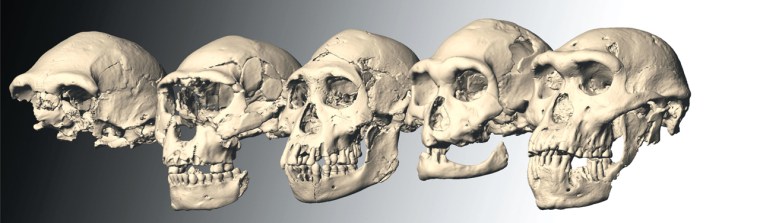

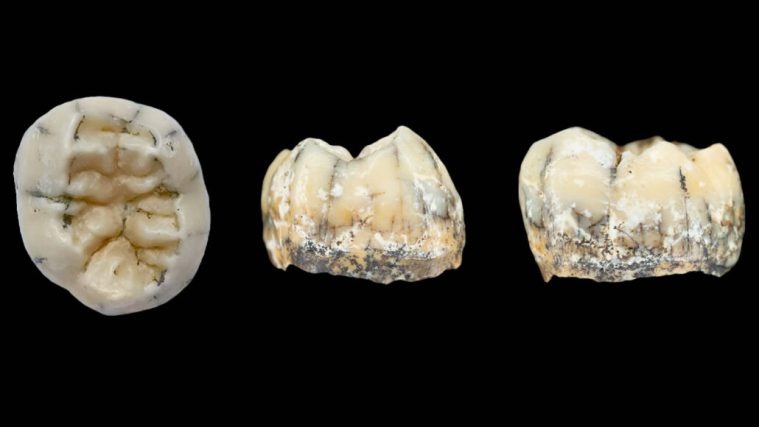
On 13 May 2026 a team led by palaeogeneticist Qiaomei Fu of the Chinese Academy of Sciences published data on proteins and amino acids yielded by enamel from six teeth of these ancient Chinese fossils, which emerged from three localities of Middle Pleistocene age (580 to 400 ka) (Fu, Q. and 11 others 2026. Enamel proteins from six Homo erectus specimens across China. Nature, v. 10.1038/s41586-026-10478-8). One is from the Zhoukoudian Cave near Beijing, famous for ‘Peking Man’, the primary reference for the anatomy of Asian H. erectus. Older fossils are unlikely to yield meaningful data of this kind because of chemical degradation; the reason why DNA has so far proved elusive from these specimens. Tooth enamel is extremely durable and can protect proteins and amino acids. Since both are produced by genes on DNA they are proxies for variants of those long-vanished genes
The key protein in the supposed H. erectus teeth isameloblastin which is involved in the formation of tooth enamel. The ameloblastin of all six teeth shared two amino acid variants; one previously unknown in other hominin lineages and perhaps unique to H. erectus, the other has been identified in Denisovans. Fu and colleagues suggest that the original bearers of the teeth – presumed to be H. erectus – had interbred with Denisovans and passed on the second variant gene. In turn that had been passed on to Asian AMH with some of whom Denisovans had interbred; remarkably 21% of living people on the Philippine archipelago carry that gene. The authors go on further to suggest that their findings support the notion that H. erectus was the source by gene-flow for ‘super-archaic’ sections of DNA found in actual Denisovan DNA from one member of that group. That is certainly a possibility, but is not the only one.
Neither the proteomics nor the morphology of the teeth, nor the anatomy of the fossils that accompany them in any way prove that they are from actual 400 ka old Homo erectus individuals. That would require at the very least protein analyses from specimens that definitely pre-date the divergence of Denisovans from Neanderthals about 600 ka ago. Remarkably, proteins have been extracted from a ~1.8 Ma old tooth yielded by the Dmanisi H. erectus site in Georgia, but that failed to reveal anything useful in this context. Maybe future work on older Chinese hominin teeth could resolve the issue. Another hypothesis is that the bearers of the analysed teeth were a population of Denisovans who themselves developed genetic variations rather than inheriting them. Proteomics is at about the same stage in its development as human genomics was before 2010 in the run-up to discovering Neanderthal and Denisovan genomes. But in the case of H. erectus the problem began with biologists’ long record of trying to simplify the natural world, especially fossils, through ‘lumping’ rather than ‘splitting’.
Humans that science has designated as different species were capable of interbreeding over tens and hundreds of thousand years, probably repeatedly and maybe habitually. That fact makes it hard to defend the concept of their speciation. There were few environments where they could not thrive, yet their migrations spread small numbers over vast areas. Continually shifting, isolated populations would diverge genetically and physically, the more so the fewer individuals were banded together. Occasionally populations would meet: an opportunity for celebration, and more, for conscious beings facing the rigours of exploration with neither territory nor resources to defend.
See also: What Homo erectus teeth from three Chinese caves tell us about who we are. Anthropology.net, 13 May 2026; Curry, A. 2026. Ghost of long-extinct ancestor lives on in people today. Science, v. 392, p. 677-678; DOI:10.1126/science.zuwthcn; Timmer, J. 2026. Protein in Homo erectus teeth suggests Denisovans gave us some of their DNA. Arstechnica, 13 May 2026; Marshall, M. 2026. Ancient teeth hint at links between Denisovans and Homo erectus. New Scientist 13 May 2026


















{kind=link}