Followers can now download newly posted annual logs for Human Evolution and Migrations covering the years 2022 to 2025. By downloading them you can get a clear idea of how palaeoanthropology has moved forward since the Covid pandemic.
Enjoy the experience if you have the time and inclination!
For decades, most of the news concerning our deep ancestry emerged from discoveries in sub-Saharan Africa at sites in Zambia, Tanzania, Kenya, South Africa, Ethiopia. The first week of 2026 decisively shifted that focus northwards to Chad and Morocco in two separate publications.
In 2002 ago the world of palaeoanthropology was in turmoil following the first discovery of fragments of what was then thought to be a hominid, or great-ape, cranium in Chad dated at around 7 Ma ago (Brunet, M. and 37 others 2002. A new hominid from the Upper Miocene of Chad, central Africa. Nature, v. 4418, p. 145-151;DOI:10.1038/nature00879). When pieced together the cranium looked like a cross between that of a chimpanzee and an australopithecine. Some suggested that the creature may have been a ‘missing link’ between the hominids and hominins; perhaps the ultimate ancestor of humans. Sahelanthropus tchadensis (nicknamedToumaï or ‘hope of life’ in the local Goran language) was undoubtedly enigmatic. The ‘molecular-clock’ age estimate for the branching of hominins from a common ancestor with chimpanzees was, in 2002, judged to be two million years later the dating of Sahelanthropus, so controversy was inevitable. Another point of contention was the size of Sahelanthropus’s canine teeth: too large for australopithecines and humans, but more appropriate for a gorilla or chimp. Moreover, Toumaï showed no indisputable evidence for having been bipedal. The Chadian site subsequently yielded three lower jaw bones and a collection of teeth, a partial femur (leg bone) and three fragmentary ulnae (forearm bones). The finds suggested that as many as five individuals had been fossilised. The femur gave an unresolved hint of an upright gait, yet the ulnas suggested Toumaï might equally have been arboreal; as could also be said for the australopithecines.
Reconstructed skull of Sahelanthropus tchadensis. (Credit: Didier Descouens, University of Toulouse)
All the limb bones of Toumaïhave now been anatomically compared with those of hominins and apes (Williams S.A. et al. 2026. Earliest evidence of hominin bipedalism in Sahelanthropus tchadensis.Science Advances, v. 12, article eadv0130; DOI: 10.1126/sciadv.adv0130). Scott Williams of New York University and co-workers from other US institutions show that although the leg bones are much the same size as those of chimpanzees, their proportions were more like those of hominins. They also showed features around the knees and hips needed for bipedalism and an insertion point for a tendon for the gluteus maximus muscle (buttock) vital for sustained upright locomotion, similar to the femurs of Orrorin tugenensis (see: Orrorin walked the walk; May 2008) and Ardipithecus ramidus. Unfortunately, an intact Sahelanthropus cranium showing a foramen magnum – where the skull attaches to the spine – continues to elude field workers. Its position distinguishes upright gait definitively.
The second new advance concerns the joint ancestry of Neanderthals, Denisovans and anatomically modern humans (AMH), whose ancient genetics crudely suggest a last common ancestor living between 765 to 550 ka. This had previously been attributed to Homo antecessor found in the Gran Dolina cave at Atapuerca in northern Spain, roughly dated between 950 ka and 770 ka. (Incidentally, Gran Dolina has yielded plausible evidence of cannibalism). A novel possibility stems from hominin fossils excavated from a cave in raised-beach sediments near Casablanca in Morocco (Hublin, JJ. and 28 others, 2026 Early hominins from Morocco basal to the Homo sapiens lineage. Nature, v. 649 ; DOI: 10.1038/s41586-025-09914-y). The fossil-bearing sediments contain evidence for a shift in the Earth’s magnetic field (the Brunhes–Matuyama reversal) dated at 773 ka, much more precisely than the Atapuerca age span for H. antecessor. Jean-Jacques Hublin of CNRS in Paris and his multinational colleagues report that the fossils are similar in age to H. antecessor, yet are morphologically distinct, displaying a combination of primitive traits and of ‘derived features reminiscent of’ later Neanderthal, Denisovan and AMH fossils. The differences and shared features suggest that there may have been genetic exchanges between the Moroccan and Iberian population over a considerable period. The most obvious route would have been across the Straits of Gibraltar, but would have required some kind of water craft. An important question is ‘which population gave rise to the other?’
Artistic reconstruction of a juvenile Homo antecessor, Based on skeletal remains from Gran Dolina Cave
Larger and more robust hominin remains in Algeria dated at 1,000 ka – H. heidelbergensis? – resemble those found near Casablanca. They may have evolved to the latter. Similar possible progenitors to Iberian Homo antecessor have yet to be found in Western Europe. Homo erectus appeared in Georgia and Romania between 2.0 and 1.9 Ma, but the intervening million years or more have yielded no credible European forebears of H. antecessor. For the moment, incursion of a North African population into Europe followed by sustained contact is Hublin et al’s favoured hypothesis, rather than a European origin for Homo antecessor. For Neanderthals and Denisovans to have originated from such an African group, as has been suggested, requires finds of African fossils with plausible resemblance to what are predominantly Eurasian groups. The Iberian population migrated far and wide in Western Europe, as witnessed by stone tools and footprints dating to between 950 to 850 ka in eastern England. So it is equally possible that the Iberian group were progenitors of Neanderthals and Denisovans in Eurasia itself. At least for the moment, ancient genomes of the two H. antecessor groups are unlikely to be found in either Iberian or African fossils of the same antiquity. But, as usual, that will not stifle debate: a resort to the adage ‘absence of evidence is not evidence of absence’ seems appropriate to several research teams!
The oldest anatomically modern human fossils dated at ~300 ka, were also discovered in Morocco (see: Origin of anatomically modern humans, June 2017). Their isolation in the NW corner of the African continent poses a similar conundrum, as since then such beings went on to occupy wide areas of sub-Saharan Africa and then the world.
As early as 3.4 Ma bones with cut marks first appear in Ethiopia suggesting that meat had by then entered the hominin diet. Access to such a rich source of protein has been suggested to have encouraged the evolution of larger hominin brains. By around2.4 Ma ago it may have led to the first known human species (Homo habilis) with a brain larger than those of australopithecines. Homo ergaster, with a significantly larger brain size, first appeared at about 1.8 Ma. As the probable inventor of bifacial stone tools and being the first hominin to leave Africa, H. ergaster needed greater cognitive abilities.It is quite likely that discovery of means to cook food then provided a further boost to human evolution. Cooking unfolds the proteins in meat and also breaks down the constituents of raw vegetables making both more palatable and easier to digest. Also, many potentially nutritious tubers are toxic if not processed and cooked. Another evolutionary advantage is that such an increased uptake of nutrients without needing an increase in successful hunting and foraging reduces the length of the ‘working day’. Fire itself provides warmth, protection from large predators and light, which further increases the time available for social and mental activities.
Being able to cook demands the controlled use of fire. But when was fire first harnessed? After 3 Ma the climate in East Africa cooled to open up dry savannah, prone to wildfires. Finding naturally roasted carcases may have been an incentive for hominins to use smouldering patches to cook meat. The next breakthrough would have been carrying embers to light fires elsewhere. The earliest tentative evidence for such a fire was discovered at Swartkrans in South Africa. Crudely dated between 2 to 1 Ma, it was a reddened patch of soil containing charred, cut marked bones and burnt biface tools. Definite evidence only appears with the burnt teeth of large carp-like fish from a 780 ka site found at Gesher Benot Ya’aqob in Israel. So, archaeological evidence for cooking is very rare. Imagine, then, the excitement of a group of archaeologists from the British Museum and the Natural History Museum in London and several universities in the UK and Netherlands at finding a small, 400 ka-old Neanderthal hearth at Barham in Kent, England containing direct evidence of how the fire was lit (Davis, R. and 14 others 2025.Earliest evidence of making fire. Nature, online advance publication. DOI: 10.1038/s41586-025-09855-6). Apparently, there are signs that fires had been lit at the hearth on twelve or more occasions. So, clearly, the spot was used regularly by Neanderthals.
Striking sparks with flint and pyrite. Credit: Craig Williams, The Trustees of the British Museum
The burnt-earth site yielded fire-cracked flint hand axes and fragments of the mineral pyrite (FeS2). Pyrite, named from the Greek word for ‘fire’ – i.e. ‘fire stone’ – creates showers of sparks when scraped with a hard, sharp tool. Struck into dry grass tinder the sparks cause it to smoulder and then burst into flame when blown on. This approach has been used throughout historic times. Interestingly, pyrite is not found in local rocks and had to have been brought from outcrops of Cretaceous Chalk 15 km away, which is also a major source of flint for stone tools. Not only had the Neanderthal ‘campers’ mastered this fire lighting method, they knew where to get the minerals required. With these skills, they could have lit fires on demand wherever they were; to cook, keep warm, light the night and keep predators at bay. The find is a lucky one, for pyrite eventually oxidises in damp air. The skill may have been acquired long before 400 ka. Yet, as most school children used to know, you can also produce fire, or at least embers and smoke(!), by abrading softwood (a schoolroom desk top) with hardwood (a wooden ruler) … Other abrasive methods are available, but none so handy as a flint-pyrite tinderbox.
See also: Smith, K.N. 2025. Getting lit: This is the oldest evidence of people starting fires. Ars Technica; 10 December 2025
One of the longest-lived hominin species that we know of was Paranthropus boisei, remains of which occur in East African sediments between 2.6 and 1.3 Ma. Others, including our own species, lasted nowhere near as long, except perhaps for Homo erectus who emerged around 1.9 Ma ago and is believed by some to have lingered on in Java until about 112 ka ago. However, when the unresolved muddle in the Middle Pleistocene of similar-looking hominin fossils is eventually unravelled – as now seems to be on the cards – that may limit the range of H. erectus to 1.9 -1.0 Ma. Paranthropoid remains are easily distinguished from those of their contemporary hominins – australopithecines and early species of Homo – being extremely robust compared with the ‘gracile’ members of the human line. They were also bipedal, but their fossil skulls are distinctive: massive teeth and jaws, and a bone crest on top of the cranium to which very powerful chewing muscles were attached. Once regarded as a sort of upright gorilla with vegetarian habits, evidence has accumulated since their first discovery that they may have been far more remarkable.
Reconstruction of a Paranthropus head (Credit: Jerry Humphrey, Pinterest)
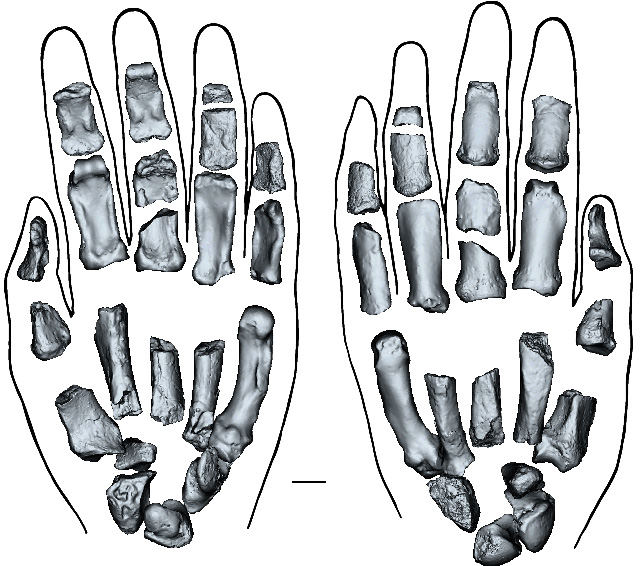
The earliest paranthropoid was P. aethiopicus from Ethiopia, dated at around 2.7 to 2.3 Ma, and believed to be the common ancestor of P. boisei and P. robustus found in Tanzania and South Africa respectively. Stone and bone tools associated with paranthropoid remains have been found in South and East Africa, some of which show signs of having been burnt. The connection between paranthropoids and both tool- and fire-making is clearly impossible to verify with certainty, and so too for their known association with australopithecine remains – or even the earliest known humans (Homo habilis) for that matter. Palaeoanthropologists are not likely to find a near-complete skeleton of any of these candidates with a tool grasped in the remains of a hand! The issue can be partly resolved if it can be shown that a fossil hand was capable of making and using tools. The fabled ‘opposable thumb’ that is capable of touching the tips of all four fingers is essential for the necessary ‘precision grip’. Isolated, 2 Ma-old thumb bones probably able to do that were found in the famous Swartkrans Cave in South Africa, but with no clue as to which hominin species had yielded them. In fact had that matter been resolved there and then, it would be not take the hominin story very far, simply because evidence for tool use – tools and cut marks on bone – goes back as far as 3.3 Ma, again with more than one candidate for the usefully endowed hominin species.
The left hand of Paranthropus boisei reconstructed from individual bones, palm-up on the left, palm down on the right. Credit: Mongle et al, Fig 3.
Remarkably, a group of scientists from the US, Canada, Australia, South Africa and Kenya have indeed unearthed from 1.5 Ma sediments on the shore of Lake Turkana in Kenya a near-complete left hand associated with cranial bones and teeth from Paranthropus boisei (Mongle,C.S. and 29 others 2025. New fossils reveal the hand of Paranthropus boisei. Nature v. 647, p. 944–951; DOI: 10.1038/s41586-025-09594-8). It is clear that the P. boisei hand shared some of the manipulative capacity of modern human hands, though it bears some resemblance to gorilla hands. That hand was probably nimble enough to make and use simple stone tools. It would also have had a powerful grip, sufficient for climbing and wielding a large stick. Yet again, it does not indicate which species first adopted tool making and use.
There are several interesting possibilities. It may be that a form of convergent evolution enabled two separate genera to become capable of such skills and the intellect to put them to use: tools, however simple, confer enormous evolutionary advantages. Had the antecedents of humans – presumably a species of Australopithecus – been the first, paranthropoids may have observed and adopted tools or vice versa. Just as possible, the – as yet unknown – common ancestor of both may have made this fundamental leap, which would have benefitted both vegetarian and omnivorous descendants. In that case the physiology of each group may have diverged with their lifestyles. Eating roots and leaves requires considerably more physical effort than getting sufficient protein and fats partly by devouring flesh.
Assigning human fossils older than around 250 ka to different groups of the genus Homo depends entirely on their physical features. That is because ancient DNA has yet to be found and analysed from specimens older than that. The phylogeny of older human remains is also generally restricted to the bones that make up their heads; 21 that are fixed together in the skull and face, plus the moveable lower jaw or mandible. Far more teeth than crania have been discovered and considerable weight is given to differences in human dentition. Teeth are not bones, but they are much more durable, having no fibrous structure and vary a great deal. The main problem for palaeoanthropologists is that living humans are very diverse in their cranial characteristics, and so it is reasonable to infer that all ancient human groups were characterised by such polymorphism, and may have overlapped in their physical appearance. A measure of this is that assigning fossils to anatomically modern humans, i.e. Homo sapiens, relies to a large extent on whether or not their lower mandible juts out to define a chin. All earlier hominins and indeed all other living apes might be regarded as ‘chinless wonders’! This pejorative term suggests dim-wittedness to most people, and anthropologists have had to inure themselves to such crude cultural conjecture.
The extraction, sequencing and comparison of ancient DNA from human fossils since 2010 has revealed that three distinct human species coexisted and interbred in Eurasia. Several well preserved examples of ancient Neanderthals and anatomically modern humans (AMH) have had their DNA sequenced, but a Denisovan genome has only emerged from a few bone fragments from the Denisova Cave in western Siberia. Whereas Neanderthals have well-known robust physical characters, until 2025 palaeoanthropologists had little idea of what Denisovans may have looked like. Then proteins and, most importantly, mitochondrial DNA (mtDNA) were extracted from a very robust skull found around 1931 in Harbin, China, dated at 146 ka. Analysis of the mtDNA and proteins, from dental plaque and bone respectively, reveal that the Harbin skull is likely to be that of a Denisovan. Previously it had been referred to as Homo longi, or ‘Dragon Man’, along with several other very robust Chinese skulls of a variety of ages.
The distorted Yunxian cranium (right) and its reconstruction (middle) [Credit: Guanghui Zhao] compared with the Harbin Denisovan cranium (left) [Hebei Geo University]
The sparse genetic data have been used to suggest the times when the three different coexisting groups diverged. DNA in Y chromosomes from Denisovans and Neanderthals suggest that the two lineages split from a common ancestor around 700 ka ago, whereas Neanderthals and modern humans diverged genetically at about 370 ka. Yet the presence of sections of DNA from both archaic groups in living humans and the discovery that a female Neanderthal from Denisova cave had a Neanderthal mother and a Denisovan father reveals that all three were interfertile when they met and interacted. Such admixture events clearly have implications for earlier humans. There are signs of at least 6 coexisting groups as far back as the Middle Pleistocene (781 to 126 ka), referred to by some as the ‘muddle in the middle’ because such an association has increasingly mystified palaeoanthropologists. A million-year-old, cranium found near Yunxian in Hubei Province, China, distorted by the pressure of sediments in which it was buried, has been digitally reconstructed.
This reconstruction encouraged a team of Chinese scientists, together with Chris Stringer of the UK Museum of Natural History, to undertake a complex statistical study of the Yunxian cranium. Their method compares it with anatomical data for all members of the genus Homo from Eurasia and Africa, i.e. as far back as the 2.4 Ma old H. habilis(Xiabo Feng and 12 others 2025. The phylogenetic position of the Yunxian cranium elucidates the origin of Homo longi and the Denisovans. Science, v. 389, p. 1320-1324; DOI: 10.1126/science.ado9202). The study has produced a plausible framework that suggests that the five large-brained humans known from 800 ka ago – Homo erectus (Asian), H. heidelbergensis, H. longi (Denisovans), H. sapiens, and H. neanderthalensis – began diverging from one another more than a million years ago. The authors regard the Yuxian specimen as an early participant in that evolutionary process. The fact that at least some remained interfertile long after the divergence began suggests that it was part of the earlier human evolutionary process. It is also possible that the repeated morphological divergence may stem from genetic drift. That process involves small populations with limited genetic diversity that are separated from other groups, perhaps by near-extinction in a population bottleneck or as a result of the founder effect when a small group splits from a larger population during migration. The global population of early humans was inevitably very low, and migrations would dilute and fragment each group’s gene pool.
The earliest evidence for migration of humans out of Africa emerged from the discovery of five 1.8 Ma old crania of H. erectus at Dmanisi to the east of the Black Sea in Georgia. similar archaic crania have been found in eastern Eurasia, especially China, at various localities with Early- to Middle Pleistocene dates. The earliest European large-brained humans – 1.2 to 0.8 Ma old H. antecessor from northern Spain – must have migrated a huge distance from either Africa or from eastern Eurasia and may have been a product of the divergence-convergence evolutionary framework suggested by Xiabo Feng and colleagues. Such a framework implies that even earlier members of what became the longi,heidelbergensis, neanderthalensis, and sapiens lineages may await either recognition or discovery elsewhere. But the whole issue raises questions about the widely held view that Homo sapiens first appeared 300 ka ago in North Africa and then populated the rest of that continent. Was that specimen a migrant from Eurasia or from elsewhere in Africa? The model suggested by Xiabo Feng and colleagues is already attracting controversy, but that is nothing new among palaeoanthropologists. Yet it is based on cutting edge phylogeny derived from physical characteristics of hominin fossils: the traditional approach by all palaeobiologists. Such disputes cannot be resolved without ancient DNA or protein assemblages. But neither is a completely hopeless task, for Siberian mammoth teeth have yielded DNA as old as 1.2 Ma and the record is held by genetic material recovered from sediments in Greenland that are up to 2.1 Ma old. The chances of pushing ancient human DNA studies back to the ‘muddle’ in the Middle Pleistocene depend on finding human fossils at high latitudes in sediments of past glacial maxima or very old permafrost, for DNA degrades more rapidly as environmental temperature rises.
Regular readers of Earth-logs will recall that the islands of Indonesia were reached by the archaic humans Homo erectus and H. floresiensis at least a million years ago. Anatomical comparison of their remains suggest that the diminutive H. floresiensis probably evolved from H. erectus under the stress of being stranded on the small, resource-poor island of Flores: a human example of island dwarfism. In fact there are anatomically modern humans (AMH) living on Flores that seem to have evolved dwarfism in the same way since AMH first arrived there between 50 and 5 ka. Incidentally, H. erectus fossils and artefacts were found by Eugene Dubois in the late 19th century at a famous site near Trinil in Java. In 2014, turned out that H. erectus had produced the earliest known art – zig-zag patterns on freshwater clam shells – between 540 and 430 ka ago. The episodic falls in global sea level due to massive accumulations of ice on land during successive Pleistocene glacial episodes aided migration by producing connections between the islands of SE Asia. They created a huge area of low-lying dryland known as ‘Sundaland’. The islands’ colonisation by H. erectus was made easy, perhaps inevitable.
The interconnection of SE Asian islands to form Sundaland (yellow) when sea level was 120 m lower than today. Even at that extreme the island of Sulawesi remained isolated by deep ocean water. Credit: based on Hakim et al Fig 1.
However, Flores and islands further east are separated from those to the west by a narrow but very deep strait. It channels powerful currents that are hazardous to small-boat crossings even today. Most palaeoanthropologists consider the colonisation of Flores by H. erectus most likely to have resulted by accident, reckoning that they were incapable of planning a crossing and building suitable craft. For AMH to have reached New Guinea and Australia around 60 ka ago, they must have developed sturdy craft and sea-faring skills. This paradigm suggests that the evolution of AMH, and thus their eventual occupation of all continents except Antarctica, must have involved a revolutionary ‘leap’ in their cognitive ability just before they left Africa. That view has been popularised by the presenter (Ella Al-Shamahi) of the 2025 BBC Television series Human – now on BBC iPlayer (requires viewers to create a free account) – in its second episode Into the Unknown. [The idea of a cognitive leap that ushered in the almost worldwide migration of anatomically modern humans was launched in 1995 by controversial anthropologist Chris Knight of University College London].
Flaked artefact, about the length of a human thumb, made of chert from excavations at Calio on Sulawesi, dated at 1.02 Ma. Credit: based on Hakim et al Fig 2
The large and peculiarly-shaped island of Sulawesi, also part of Indonesia, is notable for being the location of the earliest known figurative art; a cave painting of a Sulawesi warty pig, dated to at least 45.5 ka ago. Indonesian and Australian archaeologists working at a site near Calio in northern Sulawesi unearthed stone artefacts deep in river-terrace gravels that contain fossils of extinct pigs and dwarf elephants (Hakim, B. and 26 others 2025. Hominins on Sulawesi during the Early Pleistocene. Nature, v. 644;DOI: 10.1038/s41586-025-09348-6). The tools were struck from pebbles of hard fine-grained rocks by flaking to produce sharp edges. A combination of dating techniques – palaeomagnetism, uranium-series and electron-spin resonance – on the terrace sediments and fossils in them yielded ages ranging from 1.04 to 1.48 Ma; far older than the earliest known presence of AMH on the island (73–63 ka). The dates for an early human presence on Sulawesi tally with those from Flores. The tool makers were probably H. erectus. To reach the island from Sundaland at a time when global sea level was 120 m lower than at present would have required crossing more than 50 km of open water. It seems unlikely that such a journey could have been accidental. The migrants would have needed seaworthy craft; possibly rafts. Clearly the AMH crossings to New Guinea around 60 thousand years ago would have been far more daunting. Both land masses would have been below the horizon of any point of departure from the Indonesian archipelago, even with island ‘hopping’. Yet the Sulawesi discovery, combined with the plethora of islands both large and small, suggests that the earlier non-AMH inhabitants of Indonesia potentially could have spread further at times of very low sea level.
The colours of human skin, eyes and hair in living people across the world are determined by variants of genes (alleles) found at the same place on a chromosome. Since chromosomes are inherited from both mother and father, an individual may have the same two alleles (homozygous), or one of each (heterozygous). A dominant allele is always expressed, even if a single copy is present. A recessive allele is only expressed if the individual inherits two copies of it. Most characteristics of individuals result from the interaction of multiple genes, rather than a single gene. A commonly cited example is the coloration of eyes. If we had a single gene for eye colour – that of the iris – that had alleles just for blue (recessive or ‘b’) and one for brown (dominant or ‘B) pigmentation, brown-eyed individuals would have one or two ‘B’ alleles (bB or BB), whereas those with blue eyes would have to have two ‘blue’ alleles (bb). But inheritance is more complicated than that: there are people with green, hazel or grey eyes and even left- and right eyes of different colour. Such examples suggest that there are more than two genes affecting human eye colour, and each must have evolved as a result of mutations. Much the same goes for hair and skin coloration.
A group of scientists from the University of Ferrara in Italy have analysed highly detailed ancient DNA in anatomically modern human remains from Russia (Palaeolithic), Sweden (Mesolithic) and Croatia (Neolithic) to tease out the complexities of pigmentation inheritance. Then they applied a statistical approach learned from that study to predict the likely skin-, eye- and hair pigmentation in 348 less detailed genomes of ancient individuals whose remains date back to 45 Ma ( Silvia Perretti et al, 2025. Inference of human pigmentation from ancient DNA by genotype likelihood. Proceedings of the National Academy of Science, v. 122, article e2502158122; DOI: 10.1073/pnas.2502158122).
An artist’s impression of a Mesolithic woman from southern Denmark (credit: Tom Bjorklund)
All the hunter-gatherer Palaeolithic individuals (12 samples between 45 and 13 ka old) bar one, showed clear signs of dark pigmentation in skin, eyes and hair – the outlier from Russia was probably lighter. Those from the Mesolithic (14 to 4 ka) showed that 11 out of 35 had a light eye colour (Northern Europe, France, and Serbia), but most retained the dark skin and hair expected in descendants of migrants from Africa. Only one 12 ka hunter-gatherer from Sweden had inferred blue eyes, blonde hair, and light skin. The retention of dark pigmentation by European hunter-gatherers who migrated there from Africa has been noted before, using DNA from Mesolithic human remains and in one case from birch resin chewed by a Mesolithic woman. This called into question the hypothesis that high levels of melatonin in skin, which protects indigenous people in Africa from cancers, would result in their producing insufficient vitamin D for good health. That notion supposed that out-of-Africa migrants would quickly evolve paler skin coloration at higher latitudes. It is now known that diets rich in meat, nuts and fungi – staple for hunter-gatherers – provide sufficient vitamin-D for health at high latitudes. A more recent hypothesis is that pale skins may have evolved only after the widespread Neolithic adoption of farming when people came to rely on a diet dominated by cereals that are a poor source of vitamin-D.
However, 132 Neolithic farmers (10 to 4 ka ago) individuals studied by Perretti et al. showed increased diversity in pigmentation, with more frequent light skin tones, yet dark individuals persisted, particularly in southern and eastern Europe. Hair and eye colour showed considerable variability, the earliest sign of red hair showing up in Turkey. Even Copper- and Bronze Age samples ( 113 from 7 to 3 ka) and those from Iron Age Europeans (25 from 3 to 1.7 ka ago) still indicate common retention of dark skin, eyes and hair, although the proportion of lighter pigmentation increased in some regions of Europe. Other analyses of ancient DNA have shown that the Palaeo- and Mesolithic populations of Europe were quickly outnumbered by influx of early farmers, probably from the Anatolian region of modern Turkey, during the Neolithic. The farming lifestyle seems likely to have allowed the numbers of those who practised it to rise beyond the natural environment’s ‘carrying capacity’ for hunter-gatherers. The former inhabitants of Europe may simply have been genetically absorbed within the growing population of farmers. Much the same absorption of earlier groups seems to have happened with the westward migration from the Ukrainian and Russia steppes of the Yamnaya people and culture, culminating in the start of the European Bronze Age that reached western Europe around 2.1 ka, The Yamnaya introduced metal culture, horse-drawn wheeled vehicles and possibly Indo-European language.
So the novel probabilistic approach to ancient DNA by Perretti et al. also casts doubt on the diet-based evolution of light pigmentation at high latitudes. Instead, pulses of large population movements and thus changes in European population genetics probably account for the persistence of abundant evidence for dark pigmentation throughout Europe until historic times. The ‘lightening’ of Europeans’ physiognomy seems to have been vastly more complex than previously believed. Early Europe seems to have been almost bewilderingly diverse, which make a complete mockery of modern chauvinism and racism. The present European genetic ‘melting pot’ is surprisingly similar to that of Europe’s ancient past.
For over a century Chinese scientists have been puzzling over ancient human skulls that show pronounced brow ridges. Some assigned them to Homo, others to species that they believe were unique to China. A widely held view in China was that people now living there evolved directly from them, adhering to the ‘Multiregional Evolution’ hypothesis as opposed to that of ‘Out of Africa’. However, the issue might now have been resolved. In the last few years palaeoanthropologists have begun to suspect that these fossilised crania may have been Denisovans, but none had been subject to genetic and proteomic analysis. The few from Siberia and Tibet that initially proved the existence of Denisovans were very small: just a finger bone and teeth. Out of the blue, teeth in a robust hominin mandible dredged from the Penghu Channel between Taiwan and China yielded protein sequences that matched proteomic data from Denisovan fossils in Denisova Cave and Baishiya Cave in Tibet, suggesting that Denisovans were big and roamed widely in East Asia. In 2021 a near-complete robust cranium came to light that had been found in the 1930s near Harbin in China and hidden – at the time the area was under Japanese military occupation. It emerged only when its finder revealed its location in 2018, shortly before his death. It was provisionally called Homo longi or ‘Dragon Man’. Qiaomei Fu of the Institute of Vertebrate Paleontology and Paleoanthropology in Beijing and her colleagues have made a comprehensive study of the fossil.
The cranium found near Harbin, China belonged to a Denisovan. Credit: Hebei Geo University
It is at least 146 ka old, probably too young to have been H.erectus, but predates the earliest anatomically modern humans to have reached East Asia from Africa (~60 ka ago). The Chinese scientists have developed protein- and DNA extraction techniques akin to those pioneered at the Max Planck Institute for Evolutionary Anthropology in Leipzig. It proved impossible to extract sufficient ancient nuclear DNA from the cranium bone for definitive genomic data to be extracted, but dental plaque (calculus) adhering around the only surviving molar in the upper jaw did yield mitochondrial DNA. The mtDNA matched that found in Siberian Denisovan remains (Qiaomei Fu et al. 2025. Denisovan mitochondrial DNA from dental calculus of the >146,000-year-old Harbin cranium. Cell, v. 188, p. 1–8; DOI: 10.1016/j.cell.2025.05.040). The bone did yield 92 proteins and 122 single amino acid polymorphisms, as well as more than 20 thousand peptides (Qiaomei Fu and 8 others 2025. The proteome of the late Middle Pleistocene Harbin individual. Science, v. 388: DOI: 10.1126/science.adu9677). Again, these established a molecular link with the already known Denisovans, specifically with one of the Denisova Cave specimens. Without the painstaking research of the Chinese team, Denisovans would have been merely a genome and a proteome without much sign of a body! From the massive skull it is clear that they were indeed big people with brains much the same size as those of living people. Estimates based on the Harbin cranium suggest an individual weighing around 100 kg (220 lb or ~15 stone): a real heavyweight or rugby prop!
The work of Qiaomei Fu and her colleagues, plus the earlier, more limited studies by Tsutaya et al., opens a new phase in palaeoanthropology. Denisovans now have a genome and well-preserved parts of an entire head, which may allow the plethora of ancient skulls from China to be anatomically assigned to the species. Moreover, by extracting DNA from dental plaque for the first time they have opened a new route to obtaining genomic material: dental calculus is very much tougher and less porous than bone.
During the Middle Palaeolithic (250 to 30 ka) anatomically modern humans (AMH) and Neanderthals were engaged in new technological developments in Europe and Africa as well as in migration and social interaction. This is reflected in the tools that they left at occupation sites and the fact that most living non-Africans carry Neanderthal DNA. One of the major cultural developments was a novel means of manufacturing stone implements. It developed from the Levallois technique that involved knapping sharp-edged flakes of hard rock from larger blocks or cores. A type of tool first found at a Neanderthal site near La Quina in France is a thick flake of stone with a broad, sharp edge that shows evidence of having been resharpened many times. Most other flake tools seem to have been ‘one-offs’ that were discarded after brief usage. The Quina version was not only durable but seems to have been multipurpose. Analysis of wear patterns on the sharpened edges suggest that they were deployed in carving wood and bone, removing fat and hair from animal hides, and butchery. Such scrapers have been found over a wide area of Europe, the Middle East and NE Asia mostly at Neanderthal sites, including the famous Denisova Cave of southern Siberia that yielded the first Denisovan DNA as well as that of Neanderthals.
Making a typical Quina scraper and related tools. The toolmaker would flake pieces of stone off the core and then carefully shape the Quina scraper. (Image credit: Pei-Yuan Xiao)
Until now, the early humans of East Asia were thought not to have proceeded beyond more rudimentary tools during the Middle Palaeolithic: in fact that archaeological designation hasn’t been applied there. Recent excavations at Longtan Cave in south-west China have forced a complete revision of that view (Ruan, Q.-J., et al. 2025. Quina lithic technology indicates diverse Late Pleistocene human dynamics in East Asia. Proceedings of the National Academy of Sciences, v. 122, article e2418029122; DOI: 10.1073/pnas.2418029122). The Longtan site has yielded more than fifty scrapers and the cores from which they had been struck that clearly suggest the Quina technology had been used there. They occur in cave sediments dated at between 60 and 50 ka. As yet, no human remains have been found in the same level at Longtan, although deeper levels dated at 412 ka have yielded hominin crania, mandibular fragments, and teeth, that have been suggested to be Homo erectus.
Quina type tools in East Asia may previously have been overlooked at other hominin sites in China: re-examination of archived tool collections may show they are in fact widespread. The technology could have been brought in by migrating Neanderthals, or maybe it was invented independently by local East Asian hominins. Because most living people in China carry Denisovan DNA in the genomes so perhaps that group developed the technique before interbreeding with AMH immigrants from the west. Indeed there is no reason to discard the notion that early AMH may have imported the Quina style. A lot of work lies ahead to understand this currently unique culture at Longtan Cave. However, interpretation of another discovery published shortly after that from Longtan has spectacularly ‘stolen the thunder’ of the Qina tools, and it was made in Taiwan …
Right (top) and downward (lower) views of the partial Penghu mandible. Credit: Yousuke Kaifu University of Tokyo, Japan and Chun-Hsiang Chang Tunghai University, Taichung, from Tsutaya et al. Fig. 1 (inset)Taiwan.
About 10 years ago, Taiwanese fishers trawling in the Penghu Channel between Taiwan and China were regularly finding bones in their nets. Between 70 to 10 ka and 190 to 130 ka ago much lower sea level due to continental ice cap formation exposed the Penghu seabed. Animals and humans were thus able to move between the East Asian mainland and what is now Taiwan. The bones brought to the surface included those of elephants, water buffaloes and tigers, but one was clearly a human lower jawbone (mandible). Its shape and large molar teeth are very different from modern human mandibles and molars. A multinational team from Japan, Denmark, Taiwan and Ireland has extracted proteins from the mandible to check its genetic affinities (Tsutaya, T. and 14 others 2025. A male Denisovan mandible from Pleistocene Taiwan. Science, v. 388, p. 176-180; DOI: 10.1126/science.ads3888). Where DNA has not been preserved in bones proteomics is a useful tool, especially if results are matched with other bones that have yielded both DNA and protein sequences. In the case of the Penghu mandible, proteins from its teeth matched those of Denisovans from the Denisova Cave in Siberia which famously yielded the genome of this elusive human group. They also matched proteins from a rib found in Tibet associated with Denisovan mitochondrial DNA in cave sediments that enclosed the bones.
The three sites (Denisova, Baishiya Cave in Tibet and Penghu Channel) that have produced plausible Denisovan specimens span a large range of latitudes and altitudes. This suggests that Denisovans were capable of successful subsistence across much of East Asia. The Penghu mandible and teeth are similar to several hominin specimens from elsewhere in China that hitherto have been attributed to H. erectus. Apart from the Denisovan type locality, most of the sites have yet to be accurately dated. Having been immersed in sea water for thousands of years isotopes used in dating have been contaminated in the Panghu specimen. It can only be guessed to have lived when the seabed from which it was recovered was dry land; i.e. between 70 to 10 ka and 190 to 130 ka. China was undoubtedly occupied by Homo erectus during the early Pleistocene, but much younger fossils have been attributed to that species by Chinese palaeoanthropologists. Could it be that they are in fact Denisovans? Maybe such people independently developed the Quina knapping technique
See also: Marwick, B. 2025. Unknown human species in East Asia used sophisticated tools at the same time Neanderthals did in Europe. Live Science, 31 March 2025; Ashworth. J. 2025. Denisovan jawbone helps to reveal appearance of ancient human species. Natural History Museum News 11 April 2025.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Hominins have been making and using stone tools since at least 3.4 Ma, as shown by cut marks on bones and stone artefacts themselves. I use the sack term ‘hominin’ because the likely makers and users of the oldest tools are either australopithecines or paranthropoids, there being no fossils designates to the genus Homo of late-Pliocene age. So it might seem un-newsworthy to report that the oldest tools deliberately made from bone are now known to occur in 1.5 Ma old sediments from the famous sedimentary sequence at Olduvai Gorge in Tanzania (de la Torre, I and 8 others2025. Systematic bone tool production at 1.5 million years ago. Nature, v. 639; DOI: 10.1038/s41586-025-08652-5). To be clear, there is abundant evidence that hominins had used bones, especially sturdy long bones, for digging perhaps, much earlier in hominin history. Again, paranthropoids have been implicated in their use. The bones found at Olduvai actually show signs of manufacture into useful objects prior to their use: they show clear signs of knapping to produce points and blades. The bones are among the sturdiest known from the Pleistocene, being from elephants and hippos. Before de la Torre and colleagues found what is essentially a bone-tool factory, it was thought that systematic use of bones in such a sophisticated manner only arose between 400 to 250 ka ago among early Homo in Europe. Sadly, fossils of whoever made the tools were not found at the site. Once again, paranthropoids as well as early Homo are known to have cohabited the area at that time.
‘Front, back and side’ views of a 1.5 Ma old tool made from an elephant humerus – its upper foreleg. The scale bar represents 5 cm. (Credit: de la Torre et al.; Fig 3a)
Bifacial Acheulean stone artefacts first appear in the rock record about 300 ka before these bone tools were made. So one idea that the authors put forward is that the same kind of stone knapping technique was transferred to the more abundant massive bones of the East African Pleistocene megafauna (in the absence or rarity of suitable blocks of stone?). But it remains unclear whether or not such tools were simply selected from very large bones smashed to get at their nutritious marrow. The first possibility implies a cultural shift, whereas the latter points simply to expedience. The authors are at pains to point out that the curious million-year gap in the record of bone tools may be ascribed either to the disappearance of bone technology or simply to archaeologists who worked elsewhere having not regarding bone fragments as the products of skills. That applies equally to earlier times, when bones were indeed used, though with not so much in the way of a ‘mental template’. As de la Torre et al. conclude ‘Future research needs to investigate whether similar bone tools were already produced in earlier times, persisted during the Acheulean and eventually evolved into Middle Pleistocene bone bifaces similar in shape, size and technology to their stone counterparts’.