Former Senior Lecturer at British Open University, research in remote sensing of arid lands, groundwater exploration, Precambrian tectonics and geochemistry
Over the last few decades improved analytical techniques have made it possible to analyse tiny mineral grains for a variety of trace elements and several isotopes. Zircons obtained directly from crushed granitic igneous rocks vary in chemistry according to the magmatic processes that generated them and their tectonic context. Elevated ratios between uranium and niobium (U/Nb) and scandium and ytterbium (Sc/Yb) are characteristic of zircons in intermediate granites. These contain 52 to 63 % SiO2 – between mafic and felsic magmas – which formed by melting of hydrated mafic crust in settings akin to modern continental arcs; i.e. in subduction zones. But such partial melting can also take place where the base of continental crust delaminates and ‘drips’ into the mantle. That process is part of what is known as stagnant lid tectonics, believed by many to have been important in the Palaeoarchaean and Hadean. Such a process would have involved nearly anhydrous conditions and thus different geochemical partitioning of elements in the magmas and minerals that crystallised from them. Exposures of crystalline continental crust become increasingly rare further back in geological time, and there are none older than 4.0 Ga – i.e. of Hadean age – with a granitic component. Consequently studying the generation of continental crust in the Hadean and the early Archaean is almost entirely dependent on ancient zircons that found their way into much younger sedimentary rocks. The most famous of these occur as detrital grains in the 3.6 Ga Jack Hills conglomerate of Western Australia. Others have been extracted from similar ~3.3 Ga sedimentary rocks in the Barberton Greenstone Belt of South Africa and Eswatini.
Cartoon of possible Hadean stagnant lid tectonics, dominated by mantle plumes. (Credit: Bédard, J.H. 2018, Fig 3B, DOI: 10.1016/j.gsf.2017.01.005)
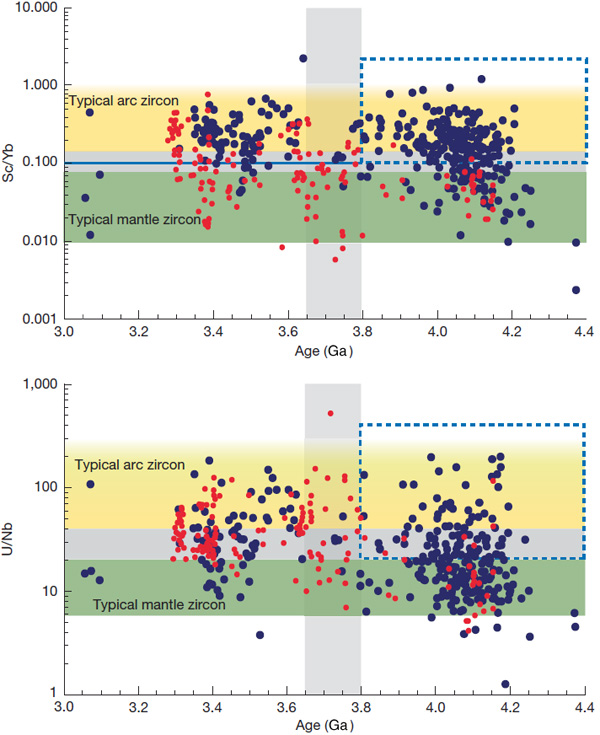
John Valley of the University of Wisconsin-Madison, USA, and co-workers from the US, Germany, Australia and France have worked on a large number of zircons newly extracted from Jack Hills. They have radiometrically dated them, and analysed Nb, Sc, U and Yb trace elements and hafnium (Hf) and oxygen isotopes Together with data from earlier studies, including Barberton zircons, they have teased out some remarkable insights into ‘continent-forming’ magmatism as far back in time as 4.4 billion years ago (Valley, J.W. and 11 others 2026. Contemporaneous mobile- and stagnant-lid tectonics on the Hadean Earth. Nature, Open access; DOI: 10.1038/s41586-025-10066-2). More than 70% of the >4.0 Ga Jack Hills zircons have elevated U/Nb and Sc/Yb ratios, which suggest that they formed in a setting akin to continental-arc subduction (CAS) zones, to produce now-vanished Hadean continental crust. The remainder seem to represent processes at mid-ocean ridge (MOR) and oceanic island (OI) settings. In contrast, the bulk of Barberton zircons of Hadean age show OI affinities, with only around 22% showing Nb–Sc–U–Yb signatures of probable CAS origins. From about 4.4 to 3.8 Ga two distinct forms of continental crust generation seem to have operated on Earth. In the erosional source region for the Barberton zircons their host granites seem to have formed during the Hadean and Eoarchaean by remelting of foundered lower crust, i.e. probably in a stagnant-lid-like tectonic setting. But at around 3.6 Ga they ‘flip’ to a subduction-like setting. The zircons yielded by Jack Hills conglomerates suggest substantially different conditions: alternating CAS and OI settings during the Hadean and a fall-off in crust generation during the Eoarchaean (4.0 to 3.8 Ga).
Plots of Sc/Yb and U/Nb against ages of zircons (vertical scale logarithmic). Black points are from Jack Hills, red from Barberton. The yellow field represents zircons formed in subduction zones; green suggests stagnant lid tectonics; grey the overlap between the two settings. Credit: Valley et al. Fig 3 a and b.
The mixed Hadean zircon signatures from Jack Hills possibly indicate that they were derived by erosion and transport from several distinct terranes that had been generated by two different processes: some kind of upper crustal recycling and stagnant lid tectonics. Meanwhile, that part of the Hadean Earth represented by the Barberton zircons may have been a long-lived regime of stagnant lid tectonics, replaced by dominant subduction at the end of the Eoarchaean. Yet the data suggest that into the Palaeoarchaean (3.6 to 3.2 Ga) and perhaps later, lid tectonics continued to operate somewhere, but at no time after 4.4 Ga was the Earth entirely subject to lid tectonics. Likewise, the authors insist that subduction was not of the plate-tectonic style, referring to some form of recycling of hydrated upper crustal mafic and ultramafic rocks into the mantle to undergo partial melting. Plate tectonics as we know it probably developed later in the Archaean. The early Earth had much higher heat flow than in later times, and thus the lithosphere was more ductile rather than brittle. The essence of modern tectonics is a series of rigid plates that extend down to the asthenosphere. When they deform it is largely through brittle failure of the entire lithosphere.
The great megalithic structure is the centrepiece of a vast ritual landscape on a 780 km2 plateau known as Salisbury Plain, underpinned by Cretaceous limestone: the largest remaining area of calcareous grassland in northwest Europe. The earliest sign that the Plain was used for ritual purposes dates to ten thousand years ago (8,000 BCE), when Mesolithic hunter gatherers erected large wooden posts to define by an E-W line the Sun’s rise and setting at the equinoxes. The area seems to have been continuously populated until 4,000 BCE when the first Neolithic farmers settled the Plain and began building burial mounds (barrows) to celebrate notable individuals and families.
The Stonehenge monument began as a circular cemetery around 3,100 BCE. Its development to the astonishing structure that remains largely intact today occupied the Neolithic populace and succeeding Bronze Age immigrants for the next 1,600 years. This involved setting up and then repeatedly shuffling around several kinds of boulders or megaliths. The first, around 2,600 BCE, were 2 to 3 tonne blocks mainly of igneous rock (the ‘bluestones’), now known to have originated from outcrops of Ordovician volcanics in Pembrokeshire about 230 km to the west. Next to arrive was a 6 tonne grey-green sandstone slab, now lying flat (hence its being named the ‘Altar’ Stone) beneath a fallen, far bigger megalith,. Once thought to be of Welsh provenance – in the Brecon Beacons 150 km to the west – the Altar Stone is now beyond a shadow of doubt to have come from Devonian strata in northern Scotland, possibly Orkney. The final erection of 30 truly enormous ‘sarsens’ to create Stonehenge’s signature circle and inner ‘horseshoe’ of vertical slabs capped by lintels took place between 2,600 to2 400 BCE. Weighing up to 50 tonnes, the sarsens are locally derived from remnants of Lower Eocene (~55 Ma) sands cemented by chemically precipitated silica (SiO2) that once covered much of southern England.
After 1,600 BCE, serious fiddling with the various stones, the bluestones in particular, ceased. The monument may have remained in some form of use during the Iron Age: it could hardly have been ignored. The first record of antiquarian interest is from the late 17th century and continued sporadically until systematic excavation of archaeological features on the Plain got underway during the 19th century and continues to the present.
Much recent literature has concentrated on what Stonehenge was for and how it was built, leading to a rich eclecticism and a little experimentation. But given the size of its stones and the obviously exotic nature of some of them, there have been disputes between those who consider them to have been brought by natural means and those who suggest collective human endeavour. The latter would have involved vast amounts of labour, shifting the bluestones over 250 km, entire community muscle power to drag the locally occurring sarsens about 25 km from their probable source, and a journey of at least 700 km to get the Altar Stone in place. Since none of the stones could conceivably have been moved by river flow, the only natural alternative for their transport is by glacial action.
Such an ice-transport theory rests on at least one of the several known advances of Pleistocene ice sheets having reached as far south as Salisbury Plain and deposited upon it glacial till that contains material from NE Scotland and South Wales. The most obvious indicators of glacial transport are large erratic boulders strewn far from their source down a previous ice stream that their distribution helps to reconstruct. In Northern Britain a great many megaliths that people erected long ago are glacial erratics of one kind or another. Of course, glacial tills contain grains of all sizes ripped and ground from the course of glacial flow. No so obvious, but equally capable of revealing transportation paths. After ice sheets melt, the till that they dump is eroded so that exotic rock and mineral grains enter drainage systems, some to remain in stream sediments. Two geologists based at Curtin University in Perth, Western Australia collected river sands from four active drainage systems on Salisbury Plain to test the glacial-transport hypothesis for the Stonehenge megaliths (Clarke, A.J.I. & Kirkland, C.L. 2026. Detrital zircon–apatite fingerprinting challenges glacial transport of Stonehenge’s megaliths. Communications Earth & Environment, v. 7, article 54; DOI: 10.1038/s43247-025-03105-3).
Using standard mineral-separation techniques – removal of low-density minerals (mainly quartz and feldspar) and those that are magnetic – Anthony Clarke and Christopher Kirkland mounted and polished samples of the remaining high-density grains embedded in resin. Using automated X-ray spectroscopy they identified grains of two minerals, zircon and apatite, that can be dated using uranium and lead isotopes. Zircons are virtually absent from the underlying Chalk although phosphorus-rich horizons in that rock sometimes contain apatite, a complex calcium phosphate. Both minerals are commonly found in igneous and metamorphic rocks and, being chemically resistant and hard, are often present in sediments derived by erosion of such parent rocks. The authors analysed U-Pb isotopes using laser ablation plasma mass spectrometry of suitable grains of each mineral. The U-Pb data from 250 apatite grains revealed a dominant age peak at 60 Ma, roughly the base of the once overlying Palaeogene sediments. Far fewer grains hint at older ages (175, 215, 300 and 625 Ma) in the Mesozoic, Palaeozoic and Neoproterozoic. The 550 analysed zircons span an age range from the Silurian to Palaeoproterozoic (432 to 1870 Ma), with a few outliers as young as 285 Ma and as old as 3396 Ma.
These data seem to suggest that they can support virtually any glacial transport hypothesis, including that of the Altar Stone, let alone the Stonehenge bluestones. However, that would be to misunderstand the complexity of sediment transport in relation to their original provenance. Erosion from a bedrock source leads to transport and deposition in sedimentary rock. Later uplift and erosion of that secondary host rock is followed by later sediment transport to another rock repository and so on and so forth through the entire geological history of Britain, across its jumble of many tectonic terranes and the effects of numerous orogenic episodes! The Salisbury Plain chalk lands were covered by Palaeogene sedimentary rocks of the London Basin. And, lo and behold, one of those younger sediments, the Thanet Formation sandstones, tell much the same U-Pb story as do the modern river sediments of Salisbury! Those Palaeocene sands elsewhere directly overlie the Chalk and, in some localities on Salisbury Plain, still do today in the form of the chemically cemented sarsens. About 50 Ma ago (early Eocene) the Palaeocene rocks and those beneath were broadly buckled by the outermost ripples of the Alpine orogeny. Once eroded from above the Plain they would certainly have delivered that signature to the mercy of subsequent back and forth river transport. And indeed the sarsens, hard to miss in that landscape, perhaps still do so. Yet no one has thought to examine their content of heavy-mineral grains.
It does seem to me that the authors, perhaps inadvertently, walked into a sedimentological minefield in a vain attempt to put the lid on the fractious debate about human- versus glacial-transport of the Stonehenge megaliths. It is not their data that fling down a ‘challenge’ to the latter hypothesis (see their Conclusions), but the widely accepted absence of even the tiniest nugget of bluestone or Devonian sandstone on the vast and heavily excavated ritual landscape of Salisbury Plain, or indeed in the gravels of the streams that currently drain the Plain. But this where the plot thickens. A recent paper by one of the proponents of the glacial hypothesis (John, B.S. 2024. A bluestone boulder at Stonehenge: implications for the glacial transport theory. E&G Quaternary Science Journal v. 73, p. 117-134;DOI: 10.5194/egqsj-73-117-2024) describes a small piece of bluestone (22 × 15 × 10 cm) that was found during excavations at Stonehenge in 1924 and mysteriously ‘rescued’ by a Robert Newall and stored in his attic for almost 50 years, eventually examined by geologists and then returned to the attic. In 1976, two years before his death Newall passed it to the curator of Salisbury Museum ‘for safe keeping’. Brian John claims that its shape and surface texture suggests glacial transport. It also has several percussion scars suggesting that it had been worked, perhaps by someone hoping to make a stone tool. Unsurprisingly, Johns succeeded in provoking a storm of criticism from archaeologists largely of the human-transport wing of the controversy. And then there is the Mumbles Erratic, found at the eponymous Mumbles headland to the west of Swansea Bay. It too looks like a ‘bluestone’, but is it an erratic or from a Neolithic ship wreck carrying bluestones from Pembrokeshire?
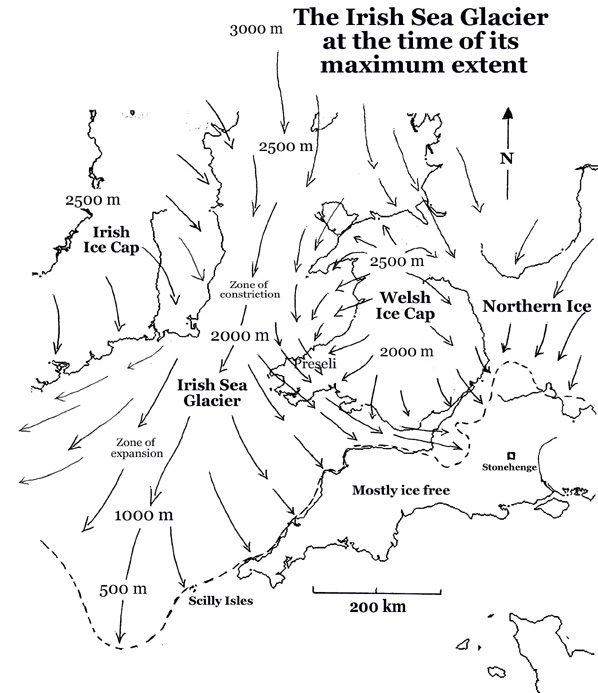
Maximum extent of glaciation in SW Britain during the Anglian Stage 478 to 424 ka ago (Credit: Wikipedia Commons)
A great deal of work by British glaciologists has established the flow patterns and extent of major ice sheets, but only for four onshore, even though there is offshore evidence for repeated glaciation back as far as 2.5 Ma ago. The most extensive of these was the Anglian Stage between 478 and 424 ka ago. The figure above shows that the Irish Sea Glacier did not reach the Stonehenge area, but it did cross Pembrokeshire to reach Somerset on the eastern side of the Bristol Channel. Bluestone erratics may have been much more easily available than blocks hewn at their source in SW Wales, an hypothesis that is currently in vogue. Nope, the quest is not over …
These days only a fool or a scoundrel would deny anthropogenic global warming and its primary outcome of inevitable sea-level rise. Yet no agency, either national or international, has set out to attempt a detailed global assessment of coastal vulnerability. There is no shortage of relevant data to do that – from remote sensing, digital elevation models, simulations of tides and wave height from meteorological data and much else. Thankfully, a team of geomorphologists, climate scientists, sociologists and economists from The Netherlands and France, led by Vindhya Basnayake of the University of Twente, The Netherlands, have taken up the challenge (Basnayake, V. et al. 2026. A global assessment of coastal vulnerability and dominant contributors. Nature Communications, in-press manuscript; DOI: 10.1038/s41467-025-67275-6).
About 10% of the world’s population – a bit less than a billion – live in coastal zones at less than 10 m elevation above mean sea level, and two-fifths that may bear the brunt of future rise. Coastal flooding and erosion threaten landforms, ecosystems and built infrastructure. Both physical effects of sea-level rise potentially may disrupt population centres, livelihoods and marine and coastal industries. More frequent and severe storms driven by global warming are also expected to increase the frequency and intensity of coastal hazards over time. Basnayake et al. have developed a Coastal Vulnerability Index (CVI) to express the hazard presented by future flooding and erosion to all coastal areas. The CVI is based on geomorphology, geology, coastal slope, coastal relief, wave height, and relative sea level change. It also integrates the local adaptive capacity and community resilience from socioeconomic and geopolitical data. Importantly CVI values are calculated at 1 km intervals along the global coastline at over 350 thousand locations. The approach used by the team incorporates from previous analyses time series for wave and tide heights and for changing sediment supply. The fine spatial resolution of data allows for identification of critical micro-regions – even within generally less vulnerable countries. Such a nuanced approach shows up the complexity of coastal risk that one-size-fits-all approaches are destined to miss.
Steep coastal slopes are less vulnerable than gentle ones, which allow greater penetration by marine hazards. The more rugged coastal terrain, the less vulnerable the coast is by acting like a large scale breakwater. Mean wave height controls the energy impinging on a coast, and is affected by wave ‘fetch’, so that ocean-facing coasts are more vulnerable than more enclosed locations. Offshore seismicity, as in island arcs, increases vulnerability to tsunamis. Tidal range has a counterintuitive effect, large ranges reducing the time a coast is in direct contact with the sea, whereas low ranges place the sea next to land for much longer. Although global sea level is destined to rise uniformly, some coasts are rising through tectonic or glacio-eustatic uplift, while others are actively subsiding; so relative sea-level change is used to address vulnerability. Other considerations assessed by Basnayake et al. are subsidence due to coastal groundwater extraction, the presence of protective coastal vegetation such as mangroves, and the influence of deltas and estuaries.
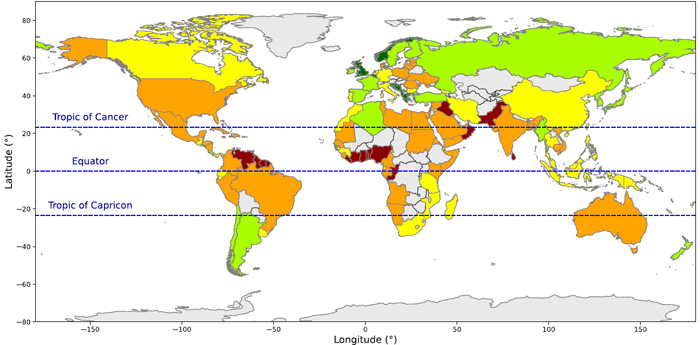
Coastal vulnerability by country: dark blue – very low; green – low; yellow – moderate; orange – high; dark red – very high. (Credit: Basnayake et al. Fig 2a)
The figure above summarises the results of the CVI study on a country-by-country basis. Eurasia is surprisingly the least vulnerable continent in this respect, especially Britain and Norway that are so exposed to the fierce North Atlantic. That is partly due to those countries high adaptive capacity and communal resilience, but mainly to their rugged and deeply indented western coasts; a legacy of glaciation. It’s important to note that coloration on the figure can be misleading. For instance, the higher resolution data pinpoint extremely high vulnerability of stretches of coast dominated by low-lying deltas, such as those of Pakistan, India, Myanmar and SE Asia. Equally surprising is the high vulnerability of North America at similar latitudes; somewhat ironic for the heartland of climate-change denial. High resolution also points to counterintuitive hazards; for instance coastal defences sometimes exacerbate vulnerability by increasing erosion on nearby undefended stretches and by hindering sediment movement. Increased onshore infrastructure boosts runoff and erosion in the coastal realm and displaces natural buffers, such as coastal forest, to storm surges: perhaps partly responsible for the high vulnerability of coasts around the hurricane belt of the Gulf of Mexico and Caribbean. Of the nineteen countries with greatest vulnerability 12 are in West Africa and NE South America and 2 in the Caribbean area. The paper is well worth reading, to get a flavour of the complexity involved and the vast magnitude of the task of ameliorating risk of coastal devastation that lies ahead in the next decades.
For decades, most of the news concerning our deep ancestry emerged from discoveries in sub-Saharan Africa at sites in Zambia, Tanzania, Kenya, South Africa, Ethiopia. The first week of 2026 decisively shifted that focus northwards to Chad and Morocco in two separate publications.
In 2002 ago the world of palaeoanthropology was in turmoil following the first discovery of fragments of what was then thought to be a hominid, or great-ape, cranium in Chad dated at around 7 Ma ago (Brunet, M. and 37 others 2002. A new hominid from the Upper Miocene of Chad, central Africa. Nature, v. 4418, p. 145-151;DOI:10.1038/nature00879). When pieced together the cranium looked like a cross between that of a chimpanzee and an australopithecine. Some suggested that the creature may have been a ‘missing link’ between the hominids and hominins; perhaps the ultimate ancestor of humans. Sahelanthropus tchadensis (nicknamedToumaï or ‘hope of life’ in the local Goran language) was undoubtedly enigmatic. The ‘molecular-clock’ age estimate for the branching of hominins from a common ancestor with chimpanzees was, in 2002, judged to be two million years later the dating of Sahelanthropus, so controversy was inevitable. Another point of contention was the size of Sahelanthropus’s canine teeth: too large for australopithecines and humans, but more appropriate for a gorilla or chimp. Moreover, Toumaï showed no indisputable evidence for having been bipedal. The Chadian site subsequently yielded three lower jaw bones and a collection of teeth, a partial femur (leg bone) and three fragmentary ulnae (forearm bones). The finds suggested that as many as five individuals had been fossilised. The femur gave an unresolved hint of an upright gait, yet the ulnas suggested Toumaï might equally have been arboreal; as could also be said for the australopithecines.
Reconstructed skull of Sahelanthropus tchadensis. (Credit: Didier Descouens, University of Toulouse)
All the limb bones of Toumaïhave now been anatomically compared with those of hominins and apes (Williams S.A. et al. 2026. Earliest evidence of hominin bipedalism in Sahelanthropus tchadensis.Science Advances, v. 12, article eadv0130; DOI: 10.1126/sciadv.adv0130). Scott Williams of New York University and co-workers from other US institutions show that although the leg bones are much the same size as those of chimpanzees, their proportions were more like those of hominins. They also showed features around the knees and hips needed for bipedalism and an insertion point for a tendon for the gluteus maximus muscle (buttock) vital for sustained upright locomotion, similar to the femurs of Orrorin tugenensis (see: Orrorin walked the walk; May 2008) and Ardipithecus ramidus. Unfortunately, an intact Sahelanthropus cranium showing a foramen magnum – where the skull attaches to the spine – continues to elude field workers. Its position distinguishes upright gait definitively.
The second new advance concerns the joint ancestry of Neanderthals, Denisovans and anatomically modern humans (AMH), whose ancient genetics crudely suggest a last common ancestor living between 765 to 550 ka. This had previously been attributed to Homo antecessor found in the Gran Dolina cave at Atapuerca in northern Spain, roughly dated between 950 ka and 770 ka. (Incidentally, Gran Dolina has yielded plausible evidence of cannibalism). A novel possibility stems from hominin fossils excavated from a cave in raised-beach sediments near Casablanca in Morocco (Hublin, JJ. and 28 others, 2026 Early hominins from Morocco basal to the Homo sapiens lineage. Nature, v. 649 ; DOI: 10.1038/s41586-025-09914-y). The fossil-bearing sediments contain evidence for a shift in the Earth’s magnetic field (the Brunhes–Matuyama reversal) dated at 773 ka, much more precisely than the Atapuerca age span for H. antecessor. Jean-Jacques Hublin of CNRS in Paris and his multinational colleagues report that the fossils are similar in age to H. antecessor, yet are morphologically distinct, displaying a combination of primitive traits and of ‘derived features reminiscent of’ later Neanderthal, Denisovan and AMH fossils. The differences and shared features suggest that there may have been genetic exchanges between the Moroccan and Iberian population over a considerable period. The most obvious route would have been across the Straits of Gibraltar, but would have required some kind of water craft. An important question is ‘which population gave rise to the other?’
Artistic reconstruction of a juvenile Homo antecessor, Based on skeletal remains from Gran Dolina Cave
Larger and more robust hominin remains in Algeria dated at 1,000 ka – H. heidelbergensis? – resemble those found near Casablanca. They may have evolved to the latter. Similar possible progenitors to Iberian Homo antecessor have yet to be found in Western Europe. Homo erectus appeared in Georgia and Romania between 2.0 and 1.9 Ma, but the intervening million years or more have yielded no credible European forebears of H. antecessor. For the moment, incursion of a North African population into Europe followed by sustained contact is Hublin et al’s favoured hypothesis, rather than a European origin for Homo antecessor. For Neanderthals and Denisovans to have originated from such an African group, as has been suggested, requires finds of African fossils with plausible resemblance to what are predominantly Eurasian groups. The Iberian population migrated far and wide in Western Europe, as witnessed by stone tools and footprints dating to between 950 to 850 ka in eastern England. So it is equally possible that the Iberian group were progenitors of Neanderthals and Denisovans in Eurasia itself. At least for the moment, ancient genomes of the two H. antecessor groups are unlikely to be found in either Iberian or African fossils of the same antiquity. But, as usual, that will not stifle debate: a resort to the adage ‘absence of evidence is not evidence of absence’ seems appropriate to several research teams!
The oldest anatomically modern human fossils dated at ~300 ka, were also discovered in Morocco (see: Origin of anatomically modern humans, June 2017). Their isolation in the NW corner of the African continent poses a similar conundrum, as since then such beings went on to occupy wide areas of sub-Saharan Africa and then the world.
I must have been about ten years old when I last saw a ‘lucky dip’ or ‘bran tub’ at a Christmas fair. You paid two shillings (now £0.1) to rootle around in the bran for 30 seconds and grab the first sizeable wrapped object that came to hand:. In my case that would be a cheap toy or trinket, but you never knew your luck as regards the top prize. There is a small asteroid called 101955 Bennu, about half a kilometre across, whose orbit around the Sun crosses that of the Earth. So it’s a bit scary, being predicted to pass within 750,000 km of Earth in September 2060 and has a 1 in 1,880 chance of colliding with us between 2178 and 2290 CE. Because Earth-crossing asteroids are a cheaper target than those in the Asteroid Belt, in 2016 NASA launched a mission named OSIRIS-REx to intercept Bennu, image it in great detail, snaffle a sample and ultimately return the sample to Earth for analysis. This wasn’t a shot in the dark, as a lot of effort and funds were expended to target and then visit Bennu. But unlike me at the fair ground, NASA will be very happy with the outcome.
The asteroid Bennu, showing its oblate spheroidal shape, due to rotation, and its rubbly structure. Source: NASA/Goddard/University of Arizona via Wikimedia Commons
Bennu is a product of what might be regarded as ‘space sedimentation’, indeed a kind of conglomerate, being made up of boulders up to 58 m across set in gravelly and finer debris or ‘regolith’. High-resolution images revealed veins of carbonate minerals in the boulders. They suggest hydrothermal activity in a much larger parent body – one of many proto-planets accreted from interstellar gas and dust as the Solar System first began to form over 4.5 billion years ago. Its collision with another sizeable body knocked off debris to send a particulate cloud towards the Sun, subsequently to clump together as Bennu by mutual gravitational attraction. The carbonate veins can only have formed by circulation of water inside Benno’s parent.
The ‘REx’ in the mission’s name is an acronym for ‘Regolith Explorer’. Sampling was accomplished on 20 October 2020 by a soft landing that drove a sample into a capsule, and then OSIRIS-REx ‘pogo-sticked’ off with the booty. The capsule was dropped off by parachute after the mission’s return on 24 September 2023, in the manner of an Amazon delivery by drone to a happy customer. So, you can understand my ‘lucky dip’ metaphor. And NASA certainly was ‘lucky’ as the contents turned out to be astonishing, as related two years later by the analytical team in the US, led by NASA’s Angel Mojarro (Mojarro, A. et al. 2025.Prebiotic organic compounds in samples of asteroid Bennu indicate heterogeneous aqueous alteration. Proceedings of the National Academy of Science, v. 122, article e2512461122; DOI: 10.1073/pnas.2512461122).
The rock itself is made from bits of carbonaceous chondrite, the most primitive matter orbiting the Sun. It contains fifteen amino acids, including all five nucleobases that make up RNA and DNA – adenine (A), guanine (G), cytosine (C), thymine (T) and uracil (U) – as in AUGC and AGCT. Benno’s complement of amino acids included 14 of the 20 used by life on Earth to synthesise proteins. The fifteenth, tryptophan, has never confidently been seen in extraterrestrial material before. Alkylated polycyclic aromatic hydrocarbons, also found in Bennu, are seen in abundance in interstellar gas clouds and comets by detecting their characteristic fluorescence when illuminated by mid-infrared radiation from hot stars using data from the Spitzer and James Webb Space Telescopes. These prebiotic organic compounds have been suggested to have played a role in the origin of life, but exposure to many produced by human activities are implicated in many cancers and cardiovascular issues. A second paper by Japanese biochemists and colleagues from the US was also published in early December 2025 (Furukawa, Y. and 13 others 2025. Bio-essential sugars in samples from asteroid Bennu. Nature Geoscience, v. 12, online article; DOI: 10.1038/s41561-025-01838). The authors identified several kinds of sugars in a sample from Bennu, including ribose – essential for building RNA – and glucose. Bennu also contains formaldehyde – a precursor of sugars – perhaps originally in the same brines in which the amino acids formed.
Yet another publication coinciding with the aforementioned two focuses on products of the oldest event in the formation of Bennu: its content of pre-solar grains (Nguyen, A.N. et al. 2025. Abundant supernova dust and heterogeneous aqueous alteration revealed by stardust in two lithologies of asteroid Bennu. Nature Astronomy, v. 9, p. 1812-1820; DOI: 10.1038/s41550-025-02688-3). In 1969 a 2 tonne carbonaceous chondrite fell near Allende in Mexico. The largest of this class ever found, it contained tiny, pale inclusions that eight years of research revealed to represent materials completely alien to the Solar System. They are characterised by proportions of isotopes of many elements that are very different from those in terrestrial materials. The anomalies could only have formed by decay of extremely short-lived isotopes that highly energetic cosmic rays produce in a manner analogous to neutron bombardment: they are products of nuclear transmutation. It is possible to estimate when the parent isotopes produced the anomalous ‘daughter’ products. One study found ages ranging from 4.6 to 7.5 Ga: up to three billion years before the Solar System began to form. It is likely that the grains are literally ‘star dust’ formed during supernovae in nearby parts of the Milky Way galaxy. Bennu samples contain six-times more presolar grains than any other chondritic meteorites. Nguyen et al. geochemically teased out grains with different nucleosynthetic origins. These ancient relics point to Bennu’s formation in a region of the presolar cloud that preceded the protoplanetary disk and was a mix of products from several stellar settings.
The results from asteroid Bennu support the key idea that that amino acid building blocks for all proteins and the nucleobases of the genetic code, together with other biologically vital compounds arose together in a primitive asteroid. Its evolution provided the physical conditions, especially the trapping of water, for the interaction of simpler components manufactured in interstellar clouds. Such ‘fertile’ planetesimals and debris from them almost certainly accreted to form planets and endowed them with the potential for life. What astonishes me is that Bennu contains the five nucleobases used in terrestrial genetics and 70% of the amino acids from which all known proteins are assembled by terrestrial life. But, as I try to explain in my book Stepping Stones: The Making of Our Home World, life as we know it arose, survived and evolved through a hugely complex concatenation of physical and chemical events lasting more than 4.5 billion years. The major events and the sequences in which they manifested themselves may indeed have been unique. Earth is a product of luck and so are we!
As early as 3.4 Ma bones with cut marks first appear in Ethiopia suggesting that meat had by then entered the hominin diet. Access to such a rich source of protein has been suggested to have encouraged the evolution of larger hominin brains. By around2.4 Ma ago it may have led to the first known human species (Homo habilis) with a brain larger than those of australopithecines. Homo ergaster, with a significantly larger brain size, first appeared at about 1.8 Ma. As the probable inventor of bifacial stone tools and being the first hominin to leave Africa, H. ergaster needed greater cognitive abilities.It is quite likely that discovery of means to cook food then provided a further boost to human evolution. Cooking unfolds the proteins in meat and also breaks down the constituents of raw vegetables making both more palatable and easier to digest. Also, many potentially nutritious tubers are toxic if not processed and cooked. Another evolutionary advantage is that such an increased uptake of nutrients without needing an increase in successful hunting and foraging reduces the length of the ‘working day’. Fire itself provides warmth, protection from large predators and light, which further increases the time available for social and mental activities.
Being able to cook demands the controlled use of fire. But when was fire first harnessed? After 3 Ma the climate in East Africa cooled to open up dry savannah, prone to wildfires. Finding naturally roasted carcases may have been an incentive for hominins to use smouldering patches to cook meat. The next breakthrough would have been carrying embers to light fires elsewhere. The earliest tentative evidence for such a fire was discovered at Swartkrans in South Africa. Crudely dated between 2 to 1 Ma, it was a reddened patch of soil containing charred, cut marked bones and burnt biface tools. Definite evidence only appears with the burnt teeth of large carp-like fish from a 780 ka site found at Gesher Benot Ya’aqob in Israel. So, archaeological evidence for cooking is very rare. Imagine, then, the excitement of a group of archaeologists from the British Museum and the Natural History Museum in London and several universities in the UK and Netherlands at finding a small, 400 ka-old Neanderthal hearth at Barham in Kent, England containing direct evidence of how the fire was lit (Davis, R. and 14 others 2025.Earliest evidence of making fire. Nature, online advance publication. DOI: 10.1038/s41586-025-09855-6). Apparently, there are signs that fires had been lit at the hearth on twelve or more occasions. So, clearly, the spot was used regularly by Neanderthals.
Striking sparks with flint and pyrite. Credit: Craig Williams, The Trustees of the British Museum
The burnt-earth site yielded fire-cracked flint hand axes and fragments of the mineral pyrite (FeS2). Pyrite, named from the Greek word for ‘fire’ – i.e. ‘fire stone’ – creates showers of sparks when scraped with a hard, sharp tool. Struck into dry grass tinder the sparks cause it to smoulder and then burst into flame when blown on. This approach has been used throughout historic times. Interestingly, pyrite is not found in local rocks and had to have been brought from outcrops of Cretaceous Chalk 15 km away, which is also a major source of flint for stone tools. Not only had the Neanderthal ‘campers’ mastered this fire lighting method, they knew where to get the minerals required. With these skills, they could have lit fires on demand wherever they were; to cook, keep warm, light the night and keep predators at bay. The find is a lucky one, for pyrite eventually oxidises in damp air. The skill may have been acquired long before 400 ka. Yet, as most school children used to know, you can also produce fire, or at least embers and smoke(!), by abrading softwood (a schoolroom desk top) with hardwood (a wooden ruler) … Other abrasive methods are available, but none so handy as a flint-pyrite tinderbox.
See also: Smith, K.N. 2025. Getting lit: This is the oldest evidence of people starting fires. Ars Technica; 10 December 2025
The emergence of the eukaryotes – of which we are a late-entry member – has been debated for quite a while. In 2023 Earth-logs reportedthat a study of ‘biomarker’ organic chemicals in Proterozoic sediments suggests that eukaryotes cannot be traced back further than about 900 Ma ago using such an approach. At about the same time another biomarker study showed signs of a eukaryote presence at around 1050 Ma. Both outcomes seriously contradicted a ‘molecular-clock’ approach based on the DNA of modern members of the Eukarya and estimates of the rate of genetic mutation. That method sought to deduce the time in the past when the last eukaryotic common ancestor (LECA) appeared. It pointed to about 2 Ga ago, i.e. a few hundred million years after the Great Oxygenation Event got underway. Since eukaryote metabolism depends on oxygen, the molecular-clock result seems reasonable. The biomarker evidence does not. But were the Palaeo- and Mesoproterozoic Eras truly ‘boring’? A recent paper by Dietmar Müller and colleagues from the Universities of Sydney and Adelaide, Australia definitely shows that geologically they were far from that (Müller, R.D. et al. 2025. Mid-Proterozoic expansion of passive margins and reduction in volcanic outgassing supported marine oxygenation and eukaryogenesis. Earth and Planetary Science Letters, v. 672; DOI: 10.1016/j.epsl.2025.119683).
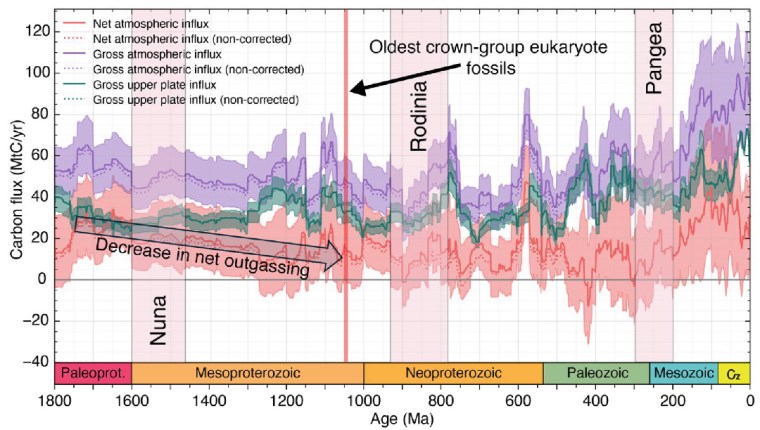
Carbon influx (million tons per year) into tectonic plates and into the ocean-atmosphere system from 1800 Ma to present. The colour bands represent: total carbon influx into the atmosphere (mauve); sequestered in tectonic plates (green); net atmospheric influx i.e. total minus carbon sequestered into plates (orange). The widths of the bands show the uncertainties of the calculated masses shown as darker coloured lines.
From 1800 to 800 Ma two supercontinents– Nuna-Columbia and Rodinia – aggregated nearly all existing continental masses, and then broke apart. Continents had collided and then split asunder to drift. So plate tectonics was very active and encompassed the entire planet, as Müller et al’s palaeogeographic animation reveals dramatically. Tectonics behaved in much the same fashion through the succeeding Neoproterozoic and Phanerozoic to build-up then fragment the more familiar supercontinent of Pangaea. Such dynamic events emit magma to form new oceanic lithosphere at oceanic rift systems and arc volcanoes above subduction zones, interspersed with plume-related large igneous provinces and they wax and wane. Inevitably, such partial melting delivered carbon dioxide to the atmosphere. Reaction on land and in the rubbly flanks of spreading ridges between new lithosphere and dissolved CO2 drew down and sequestered some of that gas in the form of solid carbonate minerals. Continental collisions raised the land surface and the pace of weathering, which also acted as a carbon sink. But they also involved metamorphism that released carbon dioxide from limestones involved in the crustal transformation. This protracted and changing tectonic evolution is completely bound up through the rock cycle with geochemical change in the carbon cycle.
From the latest knowledge of the tectonic and other factors behind the accretion and break-up of Nuna and Rodinia, Müller et al. were able to model the changes in the carbon cycle during the ‘boring billion’ and their effects on climate and the chemistry of the oceans. For instance, about 1.46 Ga ago, the total length of continental margins doubled while Nuna broke apart. That would have hugely increased the area of shallow shelf seas where living processes would have been concentrated, including the photosynthetic emission of oxygen. In an evolutionary sense this increased, diversified and separated the ecological niches in which evolution could prosper. It also increased the sequestration of greenhouse gas through reactions on the flanks of a multiplicity of oceanic rift systems, thereby cooling the planet. Translating this into a geochemical model of the changing carbon cycle (see figure) suggests that the rate of carbon addition to the atmosphere (outgassing) halved during the Mesoproterozoic. The carbon cycle and probable global cooling bound up with Nuna’s breakup ended with the start of Rodinia’s aggregation about 1000 Ma ago and the time that biomarkers first indicate the presence of eukaryotes.
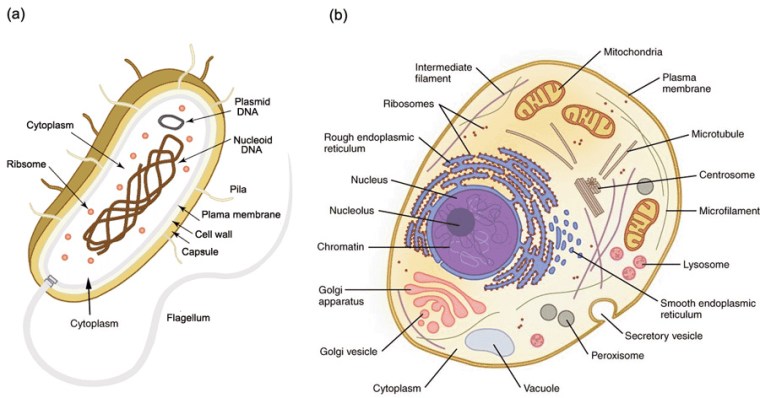
Simplified structures of (a) a prokaryote cell; (b) a simple eukaryote animal cell. Plants also contain organelles called chloroplasts
So, did tectonics play a major role in the rise of the Eukarya? Well, of course it did, as much as it was subsequently the changing background to the appearance of the Ediacaran animals and the evolutionary carnival of the Phanerozoic. But did it affect the billion-year delay of ‘eukaryogenesis’ during prolonged availability of the oxygen that such a biological revolution demanded? Possibly not. Lyn Margulis’s hypothesis of the origin of the basic eukaryote cell by a process of ‘endosymbiosis’ is still the best candidate 50 years on. She suggested that such cells were built from various forms of bacteria and archaea successively being engulfed within a cell wall to function together through symbiosis. Compared with prokaryote cells those of the eukaryotes are enormously complex. At each stage the symbionts had to be or become compatible to survive. It is highly unlikely that all components entered the relationship together. Each possible kind of cell assembly was also subject to evolutionary pressures. This clearly was a slow evolutionary process, probably only surviving from stage to stage because of the global presence of a little oxygen. But the eukaryote cell may also have been forced to restart again and again until a stable form emerged.
One of the longest-lived hominin species that we know of was Paranthropus boisei, remains of which occur in East African sediments between 2.6 and 1.3 Ma. Others, including our own species, lasted nowhere near as long, except perhaps for Homo erectus who emerged around 1.9 Ma ago and is believed by some to have lingered on in Java until about 112 ka ago. However, when the unresolved muddle in the Middle Pleistocene of similar-looking hominin fossils is eventually unravelled – as now seems to be on the cards – that may limit the range of H. erectus to 1.9 -1.0 Ma. Paranthropoid remains are easily distinguished from those of their contemporary hominins – australopithecines and early species of Homo – being extremely robust compared with the ‘gracile’ members of the human line. They were also bipedal, but their fossil skulls are distinctive: massive teeth and jaws, and a bone crest on top of the cranium to which very powerful chewing muscles were attached. Once regarded as a sort of upright gorilla with vegetarian habits, evidence has accumulated since their first discovery that they may have been far more remarkable.
Reconstruction of a Paranthropus head (Credit: Jerry Humphrey, Pinterest)
The earliest paranthropoid was P. aethiopicus from Ethiopia, dated at around 2.7 to 2.3 Ma, and believed to be the common ancestor of P. boisei and P. robustus found in Tanzania and South Africa respectively. Stone and bone tools associated with paranthropoid remains have been found in South and East Africa, some of which show signs of having been burnt. The connection between paranthropoids and both tool- and fire-making is clearly impossible to verify with certainty, and so too for their known association with australopithecine remains – or even the earliest known humans (Homo habilis) for that matter. Palaeoanthropologists are not likely to find a near-complete skeleton of any of these candidates with a tool grasped in the remains of a hand! The issue can be partly resolved if it can be shown that a fossil hand was capable of making and using tools. The fabled ‘opposable thumb’ that is capable of touching the tips of all four fingers is essential for the necessary ‘precision grip’. Isolated, 2 Ma-old thumb bones probably able to do that were found in the famous Swartkrans Cave in South Africa, but with no clue as to which hominin species had yielded them. In fact had that matter been resolved there and then, it would be not take the hominin story very far, simply because evidence for tool use – tools and cut marks on bone – goes back as far as 3.3 Ma, again with more than one candidate for the usefully endowed hominin species.
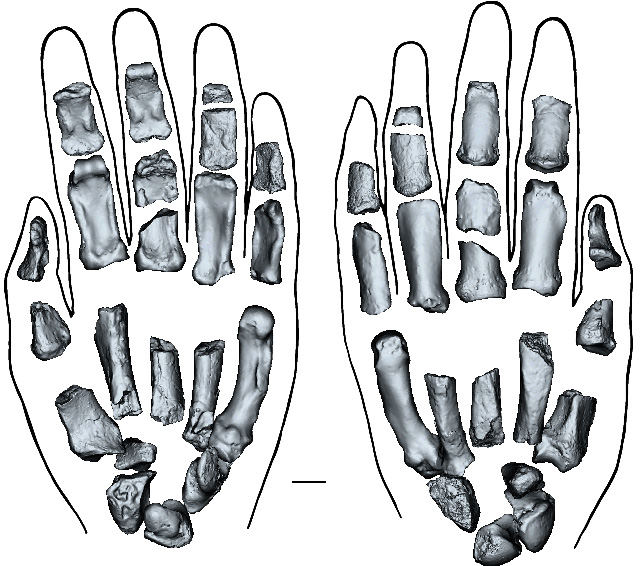
The left hand of Paranthropus boisei reconstructed from individual bones, palm-up on the left, palm down on the right. Credit: Mongle et al, Fig 3.
Remarkably, a group of scientists from the US, Canada, Australia, South Africa and Kenya have indeed unearthed from 1.5 Ma sediments on the shore of Lake Turkana in Kenya a near-complete left hand associated with cranial bones and teeth from Paranthropus boisei (Mongle,C.S. and 29 others 2025. New fossils reveal the hand of Paranthropus boisei. Nature v. 647, p. 944–951; DOI: 10.1038/s41586-025-09594-8). It is clear that the P. boisei hand shared some of the manipulative capacity of modern human hands, though it bears some resemblance to gorilla hands. That hand was probably nimble enough to make and use simple stone tools. It would also have had a powerful grip, sufficient for climbing and wielding a large stick. Yet again, it does not indicate which species first adopted tool making and use.
There are several interesting possibilities. It may be that a form of convergent evolution enabled two separate genera to become capable of such skills and the intellect to put them to use: tools, however simple, confer enormous evolutionary advantages. Had the antecedents of humans – presumably a species of Australopithecus – been the first, paranthropoids may have observed and adopted tools or vice versa. Just as possible, the – as yet unknown – common ancestor of both may have made this fundamental leap, which would have benefitted both vegetarian and omnivorous descendants. In that case the physiology of each group may have diverged with their lifestyles. Eating roots and leaves requires considerably more physical effort than getting sufficient protein and fats partly by devouring flesh.
Evidence for the earliest life on Earth has largely relied on finding signs of structures that may have been created during the Archaean Eon by micro-organisms. Actual fossils don’t turn up until the Proterozoic. The most distinctive and diverse of these are members of the Ediacaran fauna dated at around 635 Ma . The oldest widely accepted multi-celled eukaryote fossil was found in 2.1 billion-year old sediments from Gabon (see:The earliest multicelled life; July 2010). There have been a few claims for biogenic material, such as microscopic tubular structures in 3.5 billion-year (Ga) old pillow lavas and 3.2 Ga cherts from South Africa (see: Early biomarkers in South African pillow lavas; April 2004 and Believable Archaean fossils; March 2010) which some researchers dispute. Then there are Archaean stromatolites, which may be evidence for bacterial mats. The oldest of them have been claimed to occur in the famous, 3.77 Ga Isua metasediments of West Greenland. But such early fossils are chance finds, so geochemists have entered the arena with attempts to find irrefutable chemical signatures for life in ancient rocks.
One approach is isotope geochemistry. Carbon isotope data have been widely used, because life processes, such as photosynthesis, result in a deficiency of 13C relative to 12C. This was tried on graphite crystals trapped in sedimentary phosphate minerals from Isua. The results were at first acclaimed as a sign of life at around 3.8 Ga, but then refuted. In 2015 a similar approach was applied to graphite trapped in a 4.1 Ga detrital zircon, seemingly pushing back evidence for life into the Hadean. But zircon is a mineral produced by crystallisation of magma, so the fractionation of carbon isotopes in trapped graphite seem unlikely to shed light on the earliest life. The main drawback to using carbon isotopes is because metamorphism, Fischer-Tropsch mechanisms in hydrothermal environments, and volcanic processes may be responsible for enrichment of lighter carbon isotopes relative to 13C. The relative abundance of the different isotopes of iron in Archaean sediment may give clues to the transient availability of oxygen generated by bacterial photosynthesis that would oxidise soluble Fe2+ to insoluble Fe3+. Promising results were obtained in 2013 from 3.8 Ga banded ironstones at Isua. But doubt was again raised, so the only generally accepted evidence is that of the microfossils found in hydrothermal cherts in Palaeoarchaean pillow lavas from South Africa and Western Australia and the earliest stromatolites, all around 3.4 to 3.5 Ga old. However, recent research may have opened up a more convincing route to tracking down ancient life forms –actual organic molecules that make up or are produced by organisms.
Michael Wong and co-workers at the Carnegie Institution for Science in Washington, DC, USA together with other colleagues from the US, Austria, Canada, China, Belgium, Norway, Australia, the UK and France used artificial intelligence to wade through the results of geochemical analysis of over 400 ancient and modern carbon-bearing samples. (Wong, M.I. and 28 others 2025. Organic geochemical evidence for life in Archean rocks identified by pyrolysis–GC–MS and supervised machine learning. Proceedings of the National Academy of Sciences, v. 122, article e2514534122: DOI: 10.1073/pnas.2514534122). Their objective was to track the presence of organically derived molecules as far back as possible. Their approach bears a passing resemblance to that used to build genomes of ancient fossils from broken bits of DNA that reside in them. Like DNA, bio-molecules degrade over time, but leave fragments in rocks that can be detected using pyrolysis gas chromatography and mass spectrometry. In itself PGC-MS is not especially new, but using artificial intelligence (machine learning) on a massive date set certainly is: perhaps the first major trial of AI in geology.
Percentages of samples designated as biogenic by Wong et al’s AI analysis. Credit: Wong et al, Fig 4
Their samples were not just ancient rocks going back into the Archaean as far back as 3.5 Ga, but included modern biological material, meteorites presumed to have been devoid of life since their origin in pre-solar system times and synthetic samples. Wong et al divided 272 samples with known biological affinities into 9 groups to train the AI algorithm. The analytical method breaks down organic and inorganic carbonaceous materials into fragments of molecules: the opposite of DNA sequencing. When subjected to PGC-MS each type of living organism, from bacteria to animals produces a distinct pattern of molecular fragments. The AI analysis is based on a sophisticated statistical algorithm being trained to recognise ‘debris’ from organic and inorganic carbonaceous compounds according to each sample’s geochemical ‘fingerprint’. Part of the ‘training’ was based on sediments that contain irrefutable fossil samples from as far back in time as the Mesoproterozoic (1000 Ma). Another part was based on definitely inorganic materials, such as carbonaceous meteorites. AI proved able to distinguish biological from inorganic material with a probability up to 0.9 (90%). These results suggested that older, more biologically uncertain material could be assessed.
The AI was able to distinguish general biogenic affinities from inorganic ones in samples with decreasing success going back in time: as high as 0.93 in the Phanerozoic to 0.47 in the Archaean. The oldest samples that reached the probability threshold for this distinction (0.6) were 3.3 Ga cherts from the Barberton Greenstone Belt in South Africa. Another distinction between photosynthetic and and non-photosynthetic affinities among the samples that ‘passed’ as probably biotic reached the 0.6 probability threshold at 2.5 Ga for a sample from South Africa. Non-photosynthetic, but still probably biotic samples extend as far back as 3.5 Ga in South African and Western Australian Greenstone Belts.
Although Wong et al’s preliminary exploration with their novel approach doesn’t take us beyond the current 3.4 to 3.5 Ga age for the earliest tangible suggestions of life. However, they note ‘…our sample inventory is notably lacking in ancient abiogenic samples’. This is a good indication of the promise for further progress that the approach offers. Previous research has sought intact biogenic molecules, with not a great deal of luck, over several decades. Their final conclusion is ‘…information-rich attributes of ancient organic matter, even though highly degraded and with few if any surviving biomolecules, have much to reveal about the nature and evolution of life.’ They have opened a very important avenue in palaeobiological research , as their methodology seems capable of fine tuning to all manner of pro- and eukaryote biochemical distinctions. It could even be used with extraterrestrial material, should we ever get any …
Bioturbated ‘pipe rock’ of the basal Cambrian sandstones of NW Scotland. Credit: British Geological Survey photograph P531881
About 530 Ma ago most of the basic body plans of today’s living organisms can be detected as fossils, i.e. preserved hard parts. Yet studies of trace fossils (ichnofossils) – marks left in sediments by active soft bodied creatures suggest that many modern phyla arose before the start of the Cambrian (~539 Ma), as early as 545 Ma. So the term ‘Cambrian explosion’ seems to be a bit of a misnomer on two counts: it lasted around 15 Ma and began before the Cambrian. Preceding it was the Ediacaran Period that began around 100 Ma earlier in the Neoproterozoic Era. Traces of its eponymous fauna of large soft-bodied organisms are found on all continents, but apparently none of them made it into the Phanerozoic fossil record. Another characteristic of the Ediacaran is that its sedimentary rocks – and those of earlier times – show no signs of burrowing: they are not bioturbated. That may be why the Ediacaran pancake-, bun-, bag- and pen-like lifeforms are so remarkably well preserved. But a lack of burrowing did not extend to the beginning of Cambrian times. The most likely reason why it was absent during the early Ediacaran Period is that sea-floor sediments then were devoid of oxygen so eukaryote animals could not live in them. But the presence of these large organisms showed that seawater must have been oxygenated. Now clear signs of burrowing have emerged from study of Ediacaran rocks exposed in the Yangtze Gorge of Hubei,southern China ( Zhe Chen & Yarong Liu 2025. Advent of three-dimensional sediment exploration reveals Ediacaran-Cambrian ecosystem transition. Science Advances, v. 11, article eadx9449; DOI: 10.1126/sciadv.adx9449).
Tadpole-like trace fossils from the Ediacaran Dengying Formation in the Yangtze Gorge: 5 cm scale bars. The ‘heads’ show tiny depressions suggesting that there maker probed into the sediments as well as foraging horizontally. Credit: Zhe Chen & Yarong Liu; Figs 3B and 3D
Zhe Chen and Yarong Liu of the Nanjing Institute of Geology and Palaeontology and Chinese Academy of Sciences in China examined carbonates of the upper Ediacaran Dengying Formation. This overlies the Doushantuo Formation (550 to 635 Ma), known for tiny fossils of possibly the oldest deuterostome Saccorhytus coronaries; a potential candidate for the ancestor of modern bilaterian phyla. In the Yangtze Gorge locality sediments at this level show only traces of browsing of bacterial mats on the sediment surface; i.e. 2-D feeders. The basal Dengying sediments host clear signs that organisms could then penetrate into the sediments. These 3-D feeders , would have had access to buried organic remains, hitherto unexploited by living organisms. Such animal-sediment interactions would have disturbed and diminished the living microbial mats that held the sediment surface in place, and thus began to dismantle the substrate for the typical Edicaran fauna. Similar 3-D feeders occur throughout the 11 Ma represented by the Dengying Formation to the start of the Cambrian. This beginning of bioturbation heralded a period during which the Ediacaran fauna steadily waned. It also released nutrients into deep water, and opened up new ecological niches for more advanced animals on the seabed. Dissolved oxygen could only slowly enter the sediments since atmospheric and oceanic O2 levels were low. But by the earliest Cambrian it had risen to about 5 to 10% by volume to support many other kinds of burrowing animals that could penetrate more deeply, as witnessed by the abundant sandstones that occur at the base of the Cambrian in Britain.