Modern Tibetans thrive in the rarefied air at altitudes above 4 km partly because they benefit from a genetic mutation of the gene EPAS1, which regulates haemoglobin production. Surprisingly, the segment of Tibetan’s DNA that contains the mutation matches that present in the genome of an undated Denisovan girl’s finger bone found in the eponymous Siberian cave. The geneticists who made this discovery were able to estimate that Tibetans inherited the entire segment sometime in the last 40 thousand years through interbreeding with Denisovans, who probably were able to live at high altitude too. Wherever and whenever this took place the inheritance was retained because it clearly helped those who carried it to thrive in Tibet. The same segment is present in a few percent of living Han Chinese people, which suggests their ancestors and those of the Tibetans were members of the same group some 40 ka ago, most of the Han having lost the mutation subsequently.
That inheritance would have remained somewhat mysterious while the existing evidence for the colonisation of the Tibetan Plateau suggested sometime in the Holocene, possibly by migrating early farmers. A single archaeological site at 4600 m on the Plateau has changed all that (Zhang, X.L. and 15 others 2018. The earliest human occupation of the high-altitude Tibetan Plateau 40 thousand to 30 thousand years ago. Science, v. 362, p. 1049-1051; DOI: 10.1126/science.aat8824). The dig at Nwya Devu, which lies 250 km NW of Lhasa, has yielded a sequence of sediments (dated by optically stimulated luminescence at between 45 to 18 thousand years) that contains abundant stone tools made from locally occurring slate. The oldest coincides roughly with the age of the earliest anatomically modern human migrants into northern China, so the earliest Tibetans may well have been a branch of that same group of people, as suggested by the DNA of modern Tibetan and Han people. However, skeletal remains of both humans and their prey animals are yet to emerge from Nwya Devu, which leaves open the question of who they were. Anatomically modern humans or archaic humans, such as Denisovans?
The tools do not help to identify their likely makers. Slate is easy to work and typically yields flat blades with sharp, albeit not especially durable, edges; they are disposable perhaps explaining why so many were found at Nwya Devu. None show signs of pressure flaking that typify tools made from harder, more isotropic rock, such as flint. Yet they include a variety of use-types: scrapers; awls; burins and choppers as well as blades. The lack of associated remains of prey or hearths is suggested by the authors to signify that the site was a workshop; perhaps that will change with further excavation in the area. The age range suggests regular, if not permanent, occupancy for more than 20 ka
Related articles: Gibbons, A. 2014. Tibetans inherited high-altitude gene from ancient human. Science News,2 July 2014, Zhang J-F. & Dennell, R. 2018. The last of Asia conquered by Homo sapiens. Science, v. 362, p. 992-993; DOI: 10.1126/science.aav6863.
Silcrete flake from Blombos with crosshatching drawn in red ochre. (Credit: C. Foster)
The Blombos Cave 300 km east of Cape Town is where the earliest signs of art produced by anatomically modern humans were found (see Snippets on human evolution October 2011). The most publicized was a shaped piece of ochre etched with a hashed pattern of lines (Henshilwood, C.S. et al. 2018. An abstract drawing from the 73,000-year-old levels at Blombos Cave, South Africa. Nature v. 561, online; DOI: 10.1038/s41586-018-0514-3). This and the ochre-processing workshop where it was found gave a date of about 100 ka, Now another item has hit the newsrooms; a ground piece of flinty silcrete that shows signs of being the product of knapping, on which has been drawn a similar pattern, which resembles the now ubiquitous ‘hashtag’ associated with Twitter. The level in the excavation from which it was removed gives an age of about 75 ka. Like the earlier artifact, it involved the use of ochre but in a way that has been said to be an example of drawing or painting, rather than etching. It is likely to have been produced by a sharpened piece of solid ochre, perhaps a kind of crayon
For some reason the object has been hyped as the earliest example of art and of advanced cognitive abilities. But the pattern is not as complex as that on the original etched ochre block from Blombos, or even those on a freshwater mussel from Trinil in Java that could have =been made by associated Homo erectus between 430 and 500 ka ago. This does not take the context at Blombos into account. There is ample evidence that ochre, along with charcoal and burnt seal bone, was being ground there and made into paint found in an abalone shell. It can be surmised that such paint was used for some kind of decoration that has not yet been discovered. That is quite possibly because it was used for body paint as similar materials are still widely used. Now anyone – male or female – who uses cosmetics today, be it foundation, lipstick, eye-liner and -shadow or the truly fabulous make-up used by the Kathakali performers of Kerala, takes an age to try and to decide on which of an almost imperceptible range of shades to apply. Ochres are like that, as any native Australian artist will tell you.
Lord Rama face paint in Kathakali
To me, the most likely origins of both kinds of Palaeolithic hashtag are: in the case of the ‘drawing’, checking the colour and ‘grindability’ of a sharpened piece of red ochre before use; and for the etched block, using a sharp tool to grind off small amounts from what may have been a well-used block of an especially valued hue.
A revised and updated edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Two bone fragments from the Denisova Cave – the former abode of an 18th century Russian hermit called Denis – in the Altai region of Siberia yielded ancient DNA. One matches that from previously analysed Neanderthal remains and the other a genome that could only be ascribed to a hitherto unknown ancient-human population, now known as the Denisovans. Since their discovery further analysis of both modern and ancient DNA has shown that modern humans living outside of Africa contain a few percent of DNA from both ancient-human groups. Soon after leaving Africa some of their ancestors interbred with both; indeed a 40 ka-old modern-human jaw from Romania revealed genetic evidence that the individual had a Neanderthal great-great grandparent. Their descendants spread far and wide to populate Eurasia, Australasia and the Americas. Using the ancient DNA to peer back in time suggests that Neanderthals and Denisovans diverged from a common ancestor between 470 and 380 ka, itself having split from modern-human ancestry between 770 to 550 ka. Denisovan DNA also contains evidence that its ancestry included segments that could only have come from a totally unknown hominin species. Interestingly, DNA from the Neanderthal bone fragment found at Denisova contains fragments from an anatomically modern-human.
Tourists at the entrance to Denisova Cave, Russia (credit: Wikipedia)
With such riches from tiny fragments of human bones unearthed from the Denisova Cave, it is no surprise that the team led by Svante Pääbo at the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, has subsequently analysed others that showed signs of human proteins. The latest ‘takes the biscuit’. A fragment of limb bone from someone who was at least 13 years old yielded DNA commensurate with their having been the child of a Neanderthal mother and a Denisovan father (Slon, V. and 18 others 2018. The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature, v. 560, published on-line; doi: 10.1038/s41586-018-0455-x). Their child was a girl, who has been nicknamed ‘Denny’ by the team, though ‘Denise’ might seem more appropriate. The only clues to what her father, or any Denisovan, might have looked like stem from a few teeth and a skull fragment from the cave that have yielded Denisovan DNA. The teeth are much larger and the skull fragment is thicker than those of Neanderthals, suggesting that Denisovans were distinctly bigger and more robust than even the sturdy Neanderthals.
The father came from a population related to a later Denisovan found in the cave – the first to be sequenced. This suggests long-term occupancy of the area by Denisovans. But his genome also carries traces of Neanderthal ancestry. Surprisingly, the mother is more closely related to Croatian Neanderthals, rather than to an earlier Neanderthal found in the cave. Neanderthals were clearly capable of migrating between Europe and eastern Eurasia; more than 5000 km in this case. Even though very few archaic humans have been genetically sequenced it is beginning to look as if genetic mixing between diverse hominin groups in the last half million years was common, when they actually met. A custom of marrying outside a closely related group (exogamy) has been popular throughout recorded history; indeed it makes sound genetic sense. With the tiny human population density during the Late Pleistocene, it may then have been cause for mutual celebration. As documented in Chapters 2 and 3 of David Reich’s Who We Are and How We Got Here (Oxford University Press, 2018) human origins since about 470 ka until the present chart a history of episodic migrations and genetic mixing that certainly makes nonsense of earlier ideas of ‘racial purity’ and casts doubt even on the term ‘species’ as regards members of the genus Homo.
If we are ever to discover who the Denisovans were and what they looked like, the evidence is likely to come from East Asia at latitudes where climate favours preservation of DNA. Advanced sequencing equipment and techniques are now operational in China, where suspected Denisovan remains have been found
Africa is not only a large continent, but is subdivided into many different climatic zones and ecosystems and these have changed drastically over the last 2 Ma. It is further subdivided by terrain features, such as the courses of major rivers, large plateaus, tectonic rift systems and the mountains that frequently define their flanks. Getting around Africa is not easy today, was more difficult before modern transport, and many geomorphic provinces may have been mutually inaccessible in the distant past. For instance, the Sahara Desert forms a major barrier to travellers on foot because access to surface water is non-existent except at widely spaced oases. Without boats or rafts the Nile and Congo cannot be crossed for a thousand miles or more. Migration was perhaps a very rare event outside of periods of widespread humid climates or when great environmental stress forced people either to move or perish. Despite these physical and ecological divisions and barriers palaeoanthropologists have, until recently, tended to regard the evolution of Homo sapiens and earlier human and hominin species as having occurred within single populations: a linear view forced on them by scanty fossil remains and limited methodologies. Logically, when human numbers were small Africa probably had several isolated population Physical isolation would have engendered genetic isolation in which our ancestors evolved for tens of thousand years.
Anatomically modern human (AMH) remains found at Jebel Irhoud in Morocco turned out to be 315 ka old, displacing those from Ethiopia (190 ka) as the earliest known examples of AMH. Several more archaic H. sapiens fossils have turned up in southern Africa and as far afield as the Middle East, suggesting that the early evolution of AMH was in an Africa-wide context rather than in one area – the rift system of Ethiopia and Kenya – from which a new species radiated outwards. This breadth of finds has encouraged Eleanor Scerri of Oxford University and her many international colleagues to resurrect what was once a widely discarded hypothesis; a multiregional model of modern human origins, originally proposed to have arisen from pre-sapiens groups in Eurasia by Milford Wolpoff but which was sunk once genetic connections among living humans turned out to be rooted in Africa. (Scerri, E.M.L. and 22 others 2018. Did our species evolve in subdivided populations across Africa, and why does it matter? Trends in Ecology & Evolution, v. 33, p. 582-594; (PDF) doi: 10.1016/j.tree.2018.05.005). Scerri et al’s model is sited in Africa and the paper’s authors include several leading palaeoanthropologists who once opposed multiregionalism and established the Recent African Origin hypothesis on the back of the early genetic data.
Different early AMH cranium shapes: left Jebel Irhoud, Morocco (315 ka), right Qafzeh, the Levant (85 ka) (credit: Scerri et al, 2018; Figure 1)
From region to region in Africa, the oldest AMH crania show significant differences from each other, but within a distinct combination of features that clearly distinguish us from our fossil relatives and ancestors, such as Homo heidelbergensisfrom Zimbabwe and the primitive-looking H. naledifound in a South African cave in 2015. Improved dating now shows that the Zimbabwean H. heidelbergensis andH.naledi remains are roughly the same age as the Jebel Irhoud AMH specimens. The first has long been held as the progenitor of AMH and descended from H. antecessor, perhaps the common ancestor for AMH, Neanderthals and Denisovans about 700 ka ago. The three human species cohabited Africa early in the evolutionary history of AMH. It is now abundantly clear from ancient and modern genomes that AMH, Neanderthals and Denisovans interbred in Eurasia. The proximity in time and space of earlier African AMH to two more ancient human species opens up a similar possibility earlier in the emergence of all living humans. There is evidence for that too: Yoruba people living in West Africa, whose genomes have been analysed, carry up to 8% of genetic ancestry that originated in an unidentified ancient population that was non-sapiens. At present, DNA analysis with the same high precision and information content from other living Africans has not been performed, and deterioration of ancient DNA in African climates has so far thwarted genomic studies of ancient African fossils.
The new view of our origins points to repeated hybridisation involving other coexisting human species, as well as evolution in isolation, from the outset. It continued through later times while Neanderthals and Denisovans survived. Even recent human genetic history is peppered with intermingling of a great variety of migrants passing through all the habitable continents. Another issue: In the earliest times, were cultures exchanged as well as genes? The first appearance of AMH coincides with that of a new stone technology (Levallois technique), moving away from the earlier dominance by handaxes towards more delicate, leaf-shaped points, that characterise the African Middle Stone Age. Similar techniques reached Europe with the Neanderthals. Was this an invention of the earliest AMH or a joint venture?
You can find an excellent review of these issues in the September 2018 issue of Scientific American (Wong, K. 2018. Last hominin standing. Scientific American, v. 319(3), p. 56-61) along with several other articles on human evolution.
A revised and updated edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Homo floresiensis from Liang Bua Cave, Flores, Indonesia. (Credit: Wikipedia)
In October 2004 the world’s news media headlined the discovery of fossil remains of a tiny adult human on the Indonesian island of Flores, dated at around 18 ka. At only 1 m tall, with a brain cavity around a third the size of ours, yet having used stone tools and fire she was a sensational find. Someone so tiny and with such a small brain seemed highly unlikely to some palaeoanthropologists. Others claimed she was of a different species altogether. Homo floresiensis was also challenged as a new species and attributed to some congenital cause of small stature in a modern human – H. sapiens had first colonised Flores between 50 and 35 ka. But the subsequent discovery of remains of nine more individuals revealed skeletal details that were definitely un-human, with a suggestion of greater affinity to H. erectus. Her stature even suggested to a few anthropologists that she may have descended from migrant H. habilis, previously known only from 2 Ma ago in East Africa. The issue of relatedness was partly resolved by further dating of the cave strata that entombed the ‘hobbit’ which pushed her back to between 190 to 50 ka, beyond the earliest date of modern human colonisation. Further fragmentary fossil finds in more easily dated sediments on Flores showed the earliest known H.floresiensis lived around 700 ka ago. Stone tools and butchered prey remains on the island go back to 1 Ma, when the hominin trail goes cold.
Liang Bua cave where the remains of Homo floresiensis were discovered in 2003. (credit: Wikipedia)
A plausible theory for these human’s ‘hobbit’-like size is an evolutionary process known as island dwarfism, akin to that which produced the tiny elephants (Stegodon) on which they preyed. Such dramatic size reduction may arise through the influence of stringently limited food resources on the evolution of descendants from a restricted founder population, genetically cut-off from larger, more widespread populations. It now appears that such dwarfism has also affected a modern human population living on Flores (Tucci, S and 14 others 2018. Evolutionary history and adaptation of a human pygmy population of Flores Island, Indonesia. Science, v. 361, p. 511-516; doi: 10.1126/science.aar8486). A group of people of diminished stature live within shouting distance of the Liang Bua cave in which Homo floresiensis was first discovered. On average adults in the village are about 1.45 m tall. They certainly are not relict H.floresiensis, just significantly smaller than other Indonesian people living on Flores.
Serena Tucci and colleagues analysed the DNA of 32 adult pygmies from the village of Rampasasa. They show no sign of DNA from any other archaic human population than the Neanderthal and Denisovan traces that every living person outside of Africa carries – the pygmies are not descendants of H.floresiensis and are little different from other Indonesians and the rest of us. The analysis does show, however, that their ancestors carried a mixture of DNA from East Asia and New Guinea; perhaps a result of several waves of migration between 50 and 5 ka. They also carry significantly more DNA segments that are linked to short stature than do other East Asians. This suggests natural selection favored existing genes for shortness while the pygmies’ ancestors were on Flores; in other words they display an example of island dwarfism akin to that probably explaining the ‘hobbits’. Moreover, the people of Rampasasa show signs of an evolutionary adaptation to an almost exclusively meat and seafood diet, possibly arising after they migrated to Flores and had to depend on the available fauna but little in the way of plant foods.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Since discovery in 2010 that remains of the genus Homo at Dmanisi in Georgia were about 1.85 Ma old several more instances of bones and stone tools a few hundred thousand years less than that age have turned up in China. All have been ascribed to H. erectus, although there are dissimilarities with African examples of the species and its predecessor H. ergaster. The technological breakthrough that led H. erectus/ergaster to knap the distinctive bifacial or Acheulean ‘handaxe’ was achieved at about the same time as the Dmanisi humans left Africa, yet there is no sign of such tools in eastern Asia until much later, most ancient artefacts there being of a more primitive, ‘Oldowan’ type. That is perhaps because more serviceable tools were fashioned from less durable materials than fine-grained rock that takes an edge. Maybe the skills were lost en route or the forebears of eastern Asian tool makers left Africa before the breakthrough. At any rate, the genus Homo is generally conferred on any being that had a tool-making culture, so that the presence of tools alone in a sedimentary deposit signifies that humans probably once inhabited that site. The earliest tools (3.3 Ma) from the Turkana area of Kenya were made half a million years before the first known appearance of well-documented remains of an un-named member of the genus Homo at Ledi-Geraru in Afar, Ethiopia (2.8 Ma). At sites in Olduvai, Tanzania (1.9 Ma) and Turkana, Kenya (2.1 Ma) fossils of Homo habilis are found in association with ‘Oldovan’ stone tools.
Sites where early human fossils an tools have been found. (Credit: John Kappelman, Nature 2018; doi:10.1038/d41586-018-05293-9)
The latest development in the origin and wanderings of early humans has emerged from studies of a thick deposit of windblown silt or loess that makes up the Loess Plateau (Latitude 34°N) between the Yellow and Yangtze Rivers in central-east China. The loess is divided into several sequences by thin soil horizons (palaeosols). The entire stratigraphy contains tiny grains of iron minerals whose magnetic polarity was aligned with the Earth’s magnetic field at the time of deposition. This allows periods of normal and reversed geomagnetic polarity to be detected with considerable precision. Measurements have been taken at 10 cm intervals throughout the loess, to give an unbroken record of events throughout the Pleistocene Epoch that can be matched to a dated reference called the geomagnetic polarity timescale (GPTS). Palaeoclimate researchers have been able to show that the layers of loess correspond to successive glacial stages, whereas the palaeosol represent warm interglacials, exactly as recorded in sea-floor sediment profiles A team of archaeologists from China and Britain have found primitive, Oldowan-type, artefacts in both the loess and palaeosol horizons at 17 different levels (Zhu, Z. and 10 others 2018. Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago. Nature, v. 559 advance publication online doi:10.1038/s41586-018-0299-4. See also). The artefacts are positioned at levels dated at between 1.26 to 2.12 Ma by the palaeomagnetic dating (from the Réunion to Cobb Mountain normally polarized subchrons).
Primitive stone tool (four sides shown) from the Loess Plateau of China. (Credit: Zhu et al./Nature 2018)
So, in both cool and warm conditions (34°N has cold winters today) toolmakers were regularly present in central, east China for almost 900 ka. The earliest must have made a 14 thousand km trek from tropical Africa across several climatic zones, and been physically, cognitively and technologically capable of surviving and reproducing for the one- to three-thousand years the journey must have taken (based on a dispersal rate of 5 to 15 km a year estimated from modern hunter-gatherers’ activities). Either there were repeated migrations of this scale or a pioneer population survived on or within reach of the loess steppe for hundreds of thousand years. The earliest emigrants would have been neither Homo erectus nor ergaster, for neither had evolved. Their age suggests that they may have been H. habilis, a view that has been expressed for the ancestors of the diminutive H. floresiensis known to have been present of the Indonesian island of Flores for around 700 ka. Until actual fossils are unearthed – not easy as the sequence is exposed in very steep slopes characteristic of dissected loess terrains – who the first occupants of China were remains mysterious. But one thing stands out: If early humans from that long ago could arrive, survive and prosper half a world away from their place of origin, then paleoanthropologists must consider the possibility of continual diffusion of the genus Homo away from its African origins once equipped with the ability to make tools. China may become the focus for early-human research as it became for that into the origins of birds and feathered dinosaurs.
You can read more about early humans and their evolution here.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
It is often said that the biosphere is currently undergoing species losses that may rival those of the ‘Big Five’ mass extinction, with the rate of new extinctions being estimated at about 100 times the background rate during geological time. Scientifically, this is probably a dodgy assumption for palaeobiologists simply do not have the evidence to suggest what such a ‘normal’ rate might be. The fossil record is notoriously incomplete for a whole variety of reasons largely to do with both preservation and fossil collection strategies. For instance, as today, some genera may have been very common and widespread in past times, whereas others rare and restricted to small ecological niches. The record of life is prone to huge errors so that only huge, global shifts in diversity, such as mass extinctions, can be viewed with statistical rigour; and then only with caveats. For sure, the rapid demise of species today is cause for alarm and dismay, and more taxa – mainly of smaller and more restricted groups – probably have escaped identification, and will continue to do so. In the context of growing human impacts on ecosystems across the globe extinction is an increasingly emotive topic, as witness the clamour among some geoscientists for adding a new Anthropocene Epoch to the to the Stratigraphic Column. Does that require renaming the Holocene, beginning 11,700 years ago at the end of the last Ice Age, during which agriculture began? Should its start be assigned to some event during recorded history, such as the European invasion of the Americas after 1493, the beginning of the Industrial Revolution or the explosion of the first thermonuclear weapons in the 1940s and 50s? Or did humans begin significantly to affect the biosphere once their spread from Africa started after about 130 ka ago, i.e. in the late Pleistocene? That argument may well run and run: it is foremost a scientific issue, to which rules apply. A cogent example is that of the fate of megafaunas on the major continents except Antarctica as humans migrated far and wide.
The demise of the large flightless birds of Madagascar and New Zealand form a well known case as they almost certainly followed first colonisation by humans around 200 BC and 1300 CE respectively. The megafaunas of the much larger continents of Australia and the Americas have been deemed to have been more than decimated in the same way after about 65 ka and 15 ka respectively. There are no longer giant armadillos and ground sloths in South America, mammoths ceased to roam North America, and giant wombats, marsupial predators and kangaroos only remain as bones, to name but a few. It has been argued that their extinctions stemmed from the first human migrants literally eating their way through vast terrains. Yet the vast herds of Africa seem not to have been affected in the same way, until much more recently as population grew and modern projectile weapons became widely available. That has been suggested to have resulted from co-evolution of humans and megafauna over two million years, together with instinctive caution among large African beasts, whereas the ‘naivety’ of their counterparts in the Americas and Australia doomed them to extinction. Of course, it is likely that things were a great deal more complicated in every case, as argued in a review of Late Pleistocene megafaunal extinctions by Gilbert Price of the University of Queensland, and colleagues from Australia, the US and Denmark (Price, G.J. et al. 2018. Big data little help in megafauna mysteries. Nature, v. 558, p. 23-25; doi:10.1038/d41586-018-05330-7).
The gist of Price and colleagues’ critique of meta-analyses of data – 32 since 1997 – concerning allegedly human-induced extinctions is that much of the pertinent data is either low quality or poorly understood. For starters, much of the dating is questionable, either using inaccurate and outdated methods or based on inference. For instance, fossils of some alleged victim, e.g. Australian land crocodiles (Quinkana) and giant wombats (Ramsayia), have never been dated. Moreover, dates of the last known fossils are used when they may have remained extant until more recently: wooly Eurasian mammoths were long supposed not to have survived the last glacial maximum, yet recently mammoth bones from Wrangel island were found to be as young as the second millennium BCE. In 2010 spores of the fungus Sporormiella, in sediment cores, which grows only on digested plant matter in herbivore dung, was used as a proxy for the former presence or absence of large herbivore herds. Its decline in sediments after 13 ka in North America happened to coincide roughly with the start of the North American Clovis hunter culture, which was used to show that extinctions of large herbivores were linked to human predation. Yet such fungi also live on excrement of many animals both large and small, and its preservation is affected by changes in climate and water flow. To properly link declines and extinctions in human prey animals requires concrete evidence of predation, such as cut marks on identifiable bones within middens associated with human habitation, such as hearths.
When emotion, ambition and bandwagon tendencies become associated with science, objectivity sometimes gets compromised.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Since it began in 2000 Earth-Pages News has been covering the emergent science concerning the Neanderthals. To say that it has been a fertile field would be a considerable understatement, and I am certain that it will continue to be so. The first EPN item to mention them concerned the British Channel 4 TV documentary Neanderthals. In it I quoted Steve Jones FRS, former Head of the Department of Genetics, Evolution and Environment at University College London, who is always good for a humorous take on his science, and much more besides. ‘If you met an unwashed Cro Magnon dressed in a business suit on the Underground, you would probably change seats. If you met a similarly garbed Neanderthal, you would undoubtedly change trains’. That they had the build of world-champion freestyle wrestlers, beetling brows and extremely large noses was illustrated in the programme by reconstructions based on their skeletal remains; yes, they did look dangerous. But Jones’s theory is put to the test with a suitably attired actor made up as a Neanderthal man in new two-part series on BBC2, Neanderthals – Meet Your Ancestors (follow the link to watch the first programme – you need to create an account at BBC iPlayer and claim that you do have a British TV licence). Some commuters did give the chap a slightly worried glance, but there was not a rush for the doors. Ella Al-Shamahi, one of the presenters, even commented that some of her friends may well have actively sought a date with him! He certainly did not seem out of place in cosmopolitan London, and the same might be said for a cattle auction in rural Cumbria. On the New York subway he would undoubtedly have been ignored. Steve Jones was wrong!
An actor made-up to resemble a Neanderthal man in a business suit traveling on the London Underground. (Source: screen-grab from BBC2 Neanderthals – Meet Your Ancestors)
We now know that the majority of people who live on the planet today have Neanderthal segments in their genome. So between 80 and 30 ka ago to dally with Neanderthals and vice versa was as acceptable as it might be today, if they were around; probably more so, considering the amount of their DNA that is retained 2000 generations after the last possible contact. Most of the discoveries concerning Neanderthals that EPN has covered over the last 17 years are used by the two programmes to arrive at the best concept to date of just who Neanderthals were and what they were able to do.
The first episode focuses on the use of skeletal remains to visualise male and female Neanderthals, using digital techniques of forensic face and body reconstruction. Over several hundred thousand years their physiology had adapted to the fluctuating conditions of western Asia and Europe, including repeated cold episodes that ranged from full glacials to interstadials. They had also adapted to forest ecosystems where hunting would have relied on ambushing prey. Their build was suited to bursts on sprinting, probably faster than Usain Bolt could manage at his best. But they were not built for the kind of endurance needed by evolving modern humans in Africa to chase down prey in savannah scrub until they succumb to heat exhaustion. Interestingly, the right, upper-arm bones of male Neanderthals are more robust than those on the left, muscle attachment scars revealing that in life they developed large biceps, probably to use spears in powerful upward lunges into the underside of large prey, such as mammoths and aurochs. They were close-combat experts and carry the signs of injury that every such hunt would have risked. However, the series goes beyond reconstruction, by using the power of CGI motion-capture techniques developed for modern animated films and games. Actors perform the moves, and are morphed to accommodate Neanderthal physique and probable gait in the resulting action sequences – fearsomely and convincingly realistic. Analysis of a single complete hyoid bone (the hard component of the ‘voice box’ and ‘Adam’s Apple’) digitally inserted into reconstruction of the Neanderthal neck, and digital reconstruction of their possible vocal range results in demonstration of how close to human pronunciation they could have been, albeit with a pronounced distortion of the vowel Aah but almost indistinguishable for other vowels. Their faces were capable of much the same range of expressions as ours.
The second episode, available on BBC iPlayer after Sunday 20 May 2018, focuses on the genetic relations between us and Neanderthals. We gained both advantages and sets-back. It also uses what evidence there is to investigate just how like us they were in their social and intellectual behaviour. It seems they were doing OK in the grim climatic scenario of the run-up to the last glacial maximum, but succumbed to extinction during the ten thousand years following the permanent entry of anatomically modern humans into Europe and western Asia after 40 ka. Why did that happen? Every human outside sub-Saharan Africa contains Neanderthal genes: on average about 2%. Yet each of us contains a different set, so that up to 70% of the full Neanderthal genome remains in humanity as a whole. Is it possible that they could be reconstructed? Crucially, should that ever be attempted it would pose a huge moral dilemma.
To read more on the evolutionary relationships between modern humans and Neanderthals click here.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
For over half a century the presence of crude stone tools on several SE Asian islands, such as Flores and Sulawesi in Indonesia and Luzon in the Philippines, have hinted at their colonisation by Asian Homo erectus. Hominin fossils have yet to be exhumed, outside of Flores (Homo floresiensis) and dating the earlier finds has been imprecise, but evidence continues to accumulate. As regards the Philippines, the earliest hominin fossil is a modern human toe bone dated at 66.7 ka. Another curious feature of these isolated parts of what might be termed ’Wallacea’, on broader floral and faunal grounds, is the presence in the Pleistocene fossil record of large mammals, or at least dwarfed species of megafauna found in mainland Asia. These include elephants, rhinos and deer.
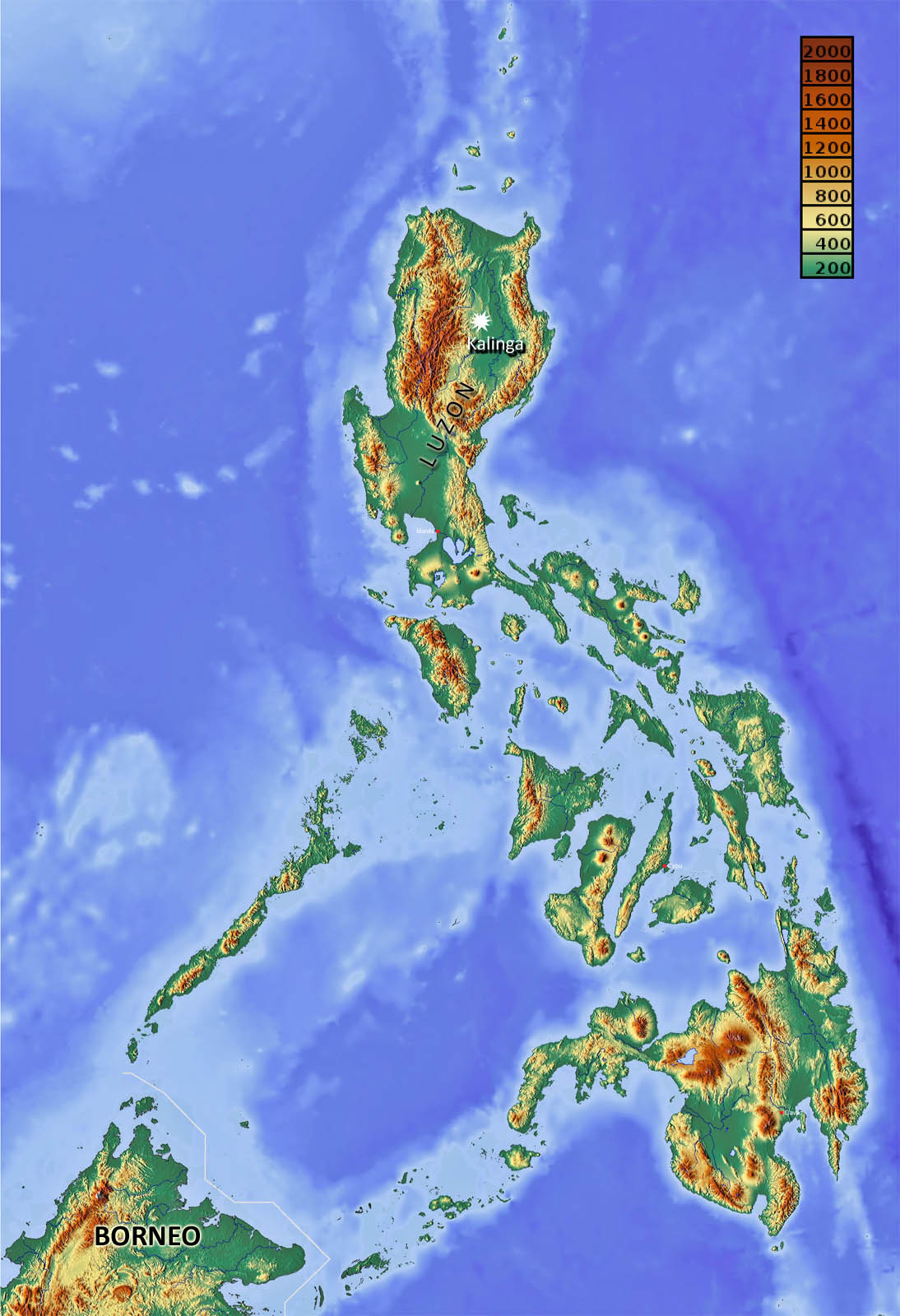
Topography of the Philippines, showing location of the Kalinga site. Palest blue sea would have been above sea level during glacial maxima. At such times Borneo would have been part of Sundaland – linked to mainland Asia (credit: Wikipedia)
A large team, with members from France, Philippines, Australia, Spain, Germany, Holland, Spain and Greece, has been excavating a tool- and fossil-rich site in thick alluvium at Kalinga in northern Luzon since 2014 (Ingicco, T. and 22 others 2018. Earliest known hominin activity in the Philippines by 709 thousand years ago. Nature, published online). It occurs in an erosional channel filled with mud. Stone artefacts comprise 56 flakes, hammer stone and cores, the tools being crude – a common feature of the Asian H. erectus lithic culture, unlike that in Africa and Europe. As well as fragments of other animals, the site is notable for a 75% complete, but disarticulated, skeleton of a rhinoceros scattered over a small area. That in itself suggests that the beast may have been butchered, and is confirmed by cut marks and signs of smashing on several of the bones. Uranium-thorium dating of one of the animal’s teeth (709±68 ka) and sediment grains from above and below the fossiliferous unit by electron-spin resonance (701±70 and 727±30 ka respectively) confirms the great antiquity of the site. Dating of volcanic plagioclase crystals from sediments by the 40Ar/39Ar method yields even older dates around 1 Ma, but the crystals may have been washed for older volcanic ash deposits.
It seems beyond doubt that early hominins, possibly H. erectus, colonised Luzon some 700 ka ago, yet, according to the authors, ‘it still seems too farfetched to suggest that H. erectus, or another unknown Pleistocene ancestral candidate … were able to construct some sort of simple watercraft and deliberately cross sea barriers’. That seems to be pushing caution a little too far. Do the authors not believe their own – to me compelling – evidence and analyses? But the paper spent a year in review, so maybe they came up against a singularly pernickety referee (three are named but one remains anonymous). Presumably, if a hominin fossil turns up during on-going excavations, that would change everything apart from the question, ‘Did they walk, swim or navigate?’ Luzon and the Philippines archipelago as a whole are surrounded by sea shallow enough for them to have been a single landmass during a glacial maximum when sea level was around 100 m lower than at present. At such a juncture a less than 20 km sea journey would have separated the Philippines from Borneo, then part of a vast area of lowland to the SW (Sundaland) that was connected to the Asian mainland.
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Ash-fall blanket at Mount Pinatubo volcano in the Philippines. (credit: Wikipedia)
The eruption that created the 100 by 30 km Toba caldera 74 ka ago was the largest recorded volcanic event during the two million years of the genus Homo’s evolution. It ejected an estimated 800 cubic kilometres of ash to blanket the land surface thousands of kilometres away. By analogy with the known effects of stratospheric ash and sulfate aerosols from the much smaller 1991 eruption of the Mount Pinatubo in the Philippines, which reduced mean global temperature by 0.5 °C, Toba might be expected to have had an even larger cooling effect, perhaps by as much as 10° C. Such a scenario has led palaeoanthropologists to suggest that there would have been major effects on humans migrating across Eurasia at the time, such as dramatic population reduction and maybe a genetic ‘bottleneck’ that could have led to rapid evolution among surviving generations. Yet, not only are stone tools found below the Toba Ash in Sumatra, but also in South India and immediately above it too. Yet analysis of Toba’s environmental effects recorded by sediments on the bed of Lake Malawi in southern African reveal little if any sign of a global ‘volcanic winter’.
A letter in Nature, published online on 15 March 2018 (Smith, E.J. and 15 others 2018. Humans thrived in South Africa through the Toba eruption about 74,000 years ago. Nature, v. 555; doi:10.1038/nature25967) decisively refutes any retardation of human cultural progress, in southern Africa at least, and suggests the opposite. Smith and colleagues from South Africa, Australia and the US found ash dated at around 74 ka 9,000 km away from Toba in sedimentary sequences that contain anatomically modern human remains and artefacts at the coastal Vleesbaai and Pinnacle Point sites in Cape Province South Africa. To check on the likelihood of a fortuitous coincidence of another eruption having shed the ash, the team compared detailed geochemical analyses of ash samples with those from other volcanoes and bona fide Toba samples, with a clear confirmation of provenance. In the archaeological record, rather than any sign of a cultural setback, the intensity of use of the sites increased after the ash-fall event, accompanied by significant technological innovations. Perhaps the Pinnacle Point community was lucky and also responded in the spirit of the adage ‘necessity is the mother of invention’. Discovery of the Toba ash at other ancient human sites would resolve the issue.
Hominin cultural revolution 320,000 years ago
As regards stone tools, the Olorgesailie Basin in southern Kenya is about as good as it gets; a long-used ‘factory’ that covers a time span from about 1. 2 to 0.03 Ma. In places, large areas of the surface underlain by sedimentary strata, some of which have become major tourist attractions, are liberally strewn with tools and debitage from their manufacture. The area is ideal for stone-tool makers: being within the East African Rift it contains outcrops of many hard, fine-grained volcanic rocks and cherts formed by hot springs interleaved with its dominant fill of lake and riverine sediments. There are two dominant sedimentary units: the Olorgesailie Formation (1.2 to 0.49 Ma) overlain by the Oltulelei Formation (0.32 to 0.05 Ma), the time gap between the two marking an extended period of regional erosion. Despite the rich tool assemblages, hominin remains have yet to be unearthed from the sediments, although there are plenty of bones from potential prey mammals. Olorgesailie was good place to live, especially as the Rift would have channelled migrating herds predictably between its steep-sided flanks.
The older formation has yielded biface tools of the Acheulean technology from bottom to top. The earliest Acheulean tools in Africa date back to about 1.7 Ma and have been attributed to Homo ergaster/erectus, although examples in Europe are associated with H. antecessor and H. heidelbergensis – the technology was active for about 1.4 Ma. The Acheulean method involved striking flakes from large blocks of rock to result in a symmetrical, pear-shaped core that served as a multipurpose tool. The oldest strata in the Oltulelei Formation contain exclusively tools that are very different , having been made by a significantly more complicated procedure and covering a wide variety of designs with different uses. This Levallois technique focused on thin flakes produced from cores after careful preparation, which enabled similar tools to be made repeatedly rather than relying on chance fracturing. Precise dating of the oldest of these assemblages gives an age of 320 ka (Deino, A.L. et al. 2018. Chronology of the Acheulean to Middle Stone Age* transition in eastern Africa. Science, v. 359 online; DOI: 10.1126/science.aao2216). The makers clearly were able to visualize the finished product within the original lump of raw stone, but in a more nuanced way than did the makers of Acheulean biface tools. The first-described Levallois tools were associated with European Neanderthals.
Producing a flake by the Levallois technique (credit: Wikipedia)
Sometime in the 500 to 320 ka interval removed by erosion a major shift in technology and almost certainly cognition took place. Not only was this a technological revolution, but the Levallois tools are found in association with a variety of pigments, such as ochres, which show signs of having been worked, presumably for decoration of some kind. Also, the tool makers seemed to have a clear preference for specific rocks – black, glassy obsidian and cherts of white and green hues from sources 25 to 90 km from the tool-making sites (Brooks, A.S. and 14 others 2018. Long-distance stone transport and pigment use in the earliest Middle Stone Age. Science, v. 359 online; DOI: 10.1126/science.aao2646). The sheer volume of tools at each site and the evidence for long-distance transport of the raw materials have prompted the authors to hazard a guess at some kind of trade, or at least cooperative intergroup interaction. Together with the use of pigment, probably for body ornamentation, this suggests individual and perhaps group identity within a kind of social network.
Of course, the big question is: Who made the leap? That’s a hard one in the absence of human remains associated with the tool-making factories at Olorgesailie . The authors of both papers argue for the earliest modern humans. But, to me, this seems like an assumption based on the age of the transition rather than any convincing evidence. The original Levallois tools from northern France were found in association with skeletal remains of Neanderthals but much later in the Pleistocene. An age of 320 ka does place the Olorgesailie tools in the same ballpark as early AMH fossils from Morocco, later than the genetically derived date of separation of Neanderthals and AMH. However, the 180 ka time gap in which the technological revolution took place gives some room for so-called African ‘archaic modern humans’ (not subdivided as are similar fossils from Europe) are known from Zimbabwe, Tanzania and Ethiopia. If the Neanderthals were using the Levallois technique in Europe there is every reason to suspect that they, or their possible forebears H. heidelbergensis, may have brought it with them from Africa.
The Olorgesailie tools figure in a third paper in the same volume of Science, but one with less shaky grounds (Potts, R. and 14 others 2018. Environmental dynamics during the onset of the Middle Stone Age in eastern Africa. Science, v. 359 online; DOI: 10.1126/science.aao2200). The wet-dry cycle of the Pleistocene, related to global warming and cooling in interglacial and glacial episodes respectively, had become more marked after the 500 to 320 ka period of tectonically induced erosion. In itself, this would have resulted in more marked shifts in the ecosystems of the basin – perhaps a case of necessity being the mother of invention. Yet, the evidence base for changing climate cycles in Africa is not from local lake-sediment stratigraphy, micropalaeontology or geochemistry, but from the modelled variation of insolation based on Milankovich’s hypothesis.
*Note: The Middle Stone Age in Africa does not correlate with the Mesolithic of Europe, but is a legacy of the development of archaeology in Africa. It corresponds to the European Middle Palaeolithic.
See also: Gibbons, A. Complex behaviour arose at dawn of humans. Science, v. 359, p. 1201-1202 (with video)
Note: Earth-Pages will be closing as of early July, but will continue in another form at Earth-logs
The first detailed description and analysis of the amazing cave paintings of Western Europe that have been attributed to anatomically modern humans (AMH) were made in the early 20th century by the Jesuit priest Abbé Henri Breuil. As well as that those of Lascaux and Altamira, which have been dated, many works in Spanish caves have not. Art ascribed to AMH includes figurative work depicting a wide range of Late Pleistocene animals, abstract and perhaps symbolic designs, and ‘signatures’ of individual people in the form of direct prints or stencils of hands. The earliest known graphic work made by modern humans is a 100 ka-old baton of ochre with a zig-zag set of sharp incisions found with ochre-filled shells possibly for body painting at Blombos Cave in South Africa.
Evidence for pre-AMH work in Europe is sparse and widely judged to be ambiguous; for instance 50 ka-old ochre-stained and pierced shells associated with Neanderthal remains in Spain. Hints at even earlier origins for art lie in the geometrically etched bivalve shells excavated by Eugene Dubois at the site in Java where he discovered Homo erectuscrania in 1891. They have recently been dated at around half a million years old. Occasionally, radiometric dating of drawings has revealed quite meagre red dots that are slightly older than the widely accepted date of first entry of AMH into Europe (~40-45 ka) and may have been made by Neanderthals. Of course, there are many European cave paintings associated with dates earlier than the extinction of Neanderthals (around 30 ka) that may have been made by them, but which are generally ascribed to AMH by assuming that only our species has the wit to make them. Even the sophisticated Châtelperronian stone tools and rough ornaments associated with undeniable Neanderthal remains are considered by many paleoanthropologists to show skills copied from AMH.
This AMH-centric view of art depends on two outlooks: simple prejudice that any beings markedly different in appearance from us were intellectually inferior – generally condemned as racist if applied to different groups of living humans; lack of incontrovertible and unambiguous evidence to the contrary. Both are set to be rigorously challenged by the growing use of sophisticated radiometric U-Th dating of the thin films of chemically precipitated calcite (flowstone or speleothem) that often coat the walls of caves and are at least as old as the art that they cover. A German-Spanish-British team has applied the technique to artwork and painted stalactites on the walls of three caves in Spain known to have been occupied by hominins over the last 100 ka (Hoffmann, D.L and 13 others 2018. U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art. Science, v. 359, p. 912-915; doi: 10.1126/science.aap7778. See also: Appenzeller, T. 2018. Europe’s first artists were Neandertals. Science, v. 359, p.852-853; doi: 10.1126/science.359.6378.852). One cave that was analysed is that at La Pasiega in Cantabria whose art was sketched by Abbé Breuil. The team’s results are dramatic: all the dated samples pre-date 40 Ka, the oldest at 79.66±14.90 ka being from La Pasiega. Precisely dated art includes hand stencils, painted stalactites, geometric patterns and line drawings of animals. Many of the caves’ artworks remain to be dated, including some well-executed animals and strange, possibly symbolic designs.
Symbolic Neanderthal art in La Pasiega cave, Spain – left: recent photograph; right: sketch produced Abbé Breuil in 1913. The red, ladder-like symbol has a minimum age of 64 ka but it is unclear if the animals and other symbols were painted later. (credit: Hoffmann et al. 2018, Supplementary Data Figure S4)
The implications of this work are far-reaching. Handprints and stencils are common throughout the archives of European cave art and seem generally to be the oldest at each site. The dating method is yet to applied to the bulk of cave art, much of which is encased in speleothem, so it is quite possible that ‘dual authorship’ may be discovered in some caves. It now seems clear that Neanderthals invented permanent art independently of AMH, and since art is a form of communication that has implications for the ability to speak as well as to think ‘outside-the-box’. The 177 ka corral-like enclosures made of stalactites and associated hearths deep within Bruniquel Cave seem more likely to have ritual significance, far from the light of day, for the Neanderthals that made them. The finds throw doubt on the implausibility of Neanderthal invention of so-called ‘transitional’ technologies, such as the Châtelperronian. Finally, fully modern humans in Africa and Neanderthals in Europe were doing much the same things over roughly the same time period; genetically and physically they parted company about 450 to 400 ka ago; both were capable of artistic symbolism and fulfilled that potential. That implies that their common ancestor may have passed on the proclivity, as might their predecessor H. erectus who created the etched mollusc shells of Trinil half a million years ago.
In June 2017 the likely age of the earliest anatomically modern humans (AMH) was pushed back to almost 300 ka with the dating of their remains found at Jebel Irhoud in Morocco. It seemed only a matter of time before their first departure from Africa would also be shown to be earlier than generally believed at between 90 to 120 ka measured from AMH remains in the Skhul and Qafzeh caves of Israel. Such an exodus may be reflected by dates (80 to 113 ka) from fragmentary and indeterminate human remains in China, but a more definite, far-travelled AMH presence in east Asia is, so far, limited to about 60 ka. Yet there is genetic evidence from Neanderthal DNA from Germany and Siberia for human-Neanderthal interbreeding at some time between 219 and 460 thousand years before present: a very hazy intimation but one that needs accounting for. The main phase of genetic introgression from Neanderthals into Homo sapiens has been estimated to have occurred at between 50 to 60 ka; more easily explained by the known AMH peregrination into Asia in that period.
Misliya Cave on Mount Carmel, Israel has now added to the Levantine AMH record. A partial upper jaw and some teeth provide morphological data that fall within the range of H. sapiens fossils, along with tools ascribed to the Levallois technology. This involved striking flakes from a prepared core – a tortoise-like bulge on the flake that detaches when struck properly to form a pre-sharpened flake, flat on one side and rounded on the other. This method was shared by both AMH and Neanderthals, and examples of the tools extend as far back as 500 ka in Africa and may have been invented by a common ancestor of both human groups. Levallois tools were found with the AMH fossils at Jebel Irhoud and also in the Levant at Tabun, dated at 190 to 260 ka, but with no associated fossil remains of their makers. Those at Mislya Cave yielded a mean age from the use of three different dating methods at least 177 ka ago, making the fossil jaw found with them the earliest direct sign of AMH outside Africa (Hershkovitz, I. and 34 others 2018. The earliest modern humans outside Africa. Science, v. 359, p. 456-459; doi: 10.1126/science.aap8369).
So, Mislya supports the genetic evidence of human-Neanderthal Introgression in Eurasia (see; Stringer, C & Galway-Witham, J. 2018. When did modern humans leave Africa? Science, v. 359, p. 389-390; doi: 10.1126/science.aas8954) and provides a spur to extend work in China and between Arabia and eastern Asia. For decades the anatomically modern human remains in the Levant have been sidelined, that near-Mediterranean area being widely regarded as a ‘boulevard of broken dreams’. That is, until Levalloisian tools dated at up to 125 ka were found in the United Arab Emirates and Arabia as a whole had been shown to have had a monsoonal climate during the glacial period that preceded the last, Eemian interglacial and in several later episodes. Once in the Levant, and provided they continually had a foothold there, AMH had many windows of opportunity to move further east without having to await falls in sea-level to open routes such as that across the Red Sea via Straits of Bab el Mandab.
Thanks to a variety of archaeological finds of tools and animal bones bearing cut marks, together with precise dating, it now seems clear that the Americas began to be colonised as early as the Last Glacial Maximum from tangible evidence from Bluefish Cave in the Yukon territory of Canada and as early as 15.5 ka close to the southern tip of South America in Chile. Although confirmation remains to be found, there is even a possibility that pre-sapiens people had arrived far earlier. Advances in analysis of ancient genetic material help understand the divergence of early colonisers. Y-chromosome DNA from living indigenous men suggests that all early Americans stemmed from 4 separate colonising populations who may have entered by crossing the Beringia land bridge, exposed as a result of glacial fall in global sea level, to follow different routes, including along the Pacific coast. A possible common ancestor of all native Americans emerged in 2013 from the mitochondrial and Y-chromosome DNA of the skeleton of a young man from near Lake Baikal in Siberia who lived about 24 ka ago. At the very start of 2018 an online paper in Nature took the story even further.
The diversity of Native American people. (credit: Wikipedia from a 19th century Norwegian painting)
The remains of a ~6-week-old girl recovered from a site at Upward Sun River in Alaska – called ‘Xach’itee’aanenh t’eede gay’, or ‘sunrise girl-child’ by indigenous Alaskans – dated at 11.5 ka, has yielded a precise genome (Moreno-Mayar, J. and 17 others 2018. Terminal Pleistocene Alaskan genome reveals first founding population of Native Americans. Nature; doi:10.1038/nature25173). The baby girl’s DNA shows that the group to which she belonged was ancestral to contemporary and fossilised ancient Native Americans. She was probably a member of a founding population of ‘Beringians’. At the end of the last glacial epoch (11.5 ka) a separate branch of Native Americans was already established in unglaciated North America further south. That group had split into two further groups sometime between 17.5 to 14.6 ka, who became ancestors of most of the indigenous people of the Americas. The ‘Beringian’ people were therefore probably stranded in the far north by the difficulties of crossing the vast North American ice sheet. Probing deeper into time, using demographic modelling, suggests that the founding population of all Native Americans, including the ‘Beringians’, split from East Asians around 36 ka ago. Gene flow among them and with East Asians persisted until about 25 thousand years ago, with some admixture with ancient northern Eurasians up to 20 ka. It seems that the ‘Beringians’, of whom little ‘sunrise girl-child’ was a late member, became isolated genetically between 22-18 ka.
The ancestral mixture of both East Asian and northern Eurasians that led to the founders of the whole panoply of geographically isolated Native Americans is remarkable. It shows just how far human groups moved and mingled during the run-up to the Last Glacial Maximum, which made the far north just about uninhabitable – or so it has been assumed. For a small ethnically mixed group to survive such conditions for so long suggests considerable ingenuity in living off the land.
When first mooted, the Out of Africa model for the spread of anatomically modern humans (AMH) centred on a single exodus from African to Eurasia, which researchers broadly agreed to have occurred about 60 thousand years ago. That was when an advance of continental glaciers and sea level fall narrowed to manageable proportions the obstacle presented by the Red Sea. The only archaeological drawback was that AMH had occupied the Levant at around 110 ka. That was formerly considered to have been a temporary occupation corralled by hyperarid conditions immediately to the east and a mountain barrier to the north, with the Mediterranean Sea to the west. Yet, during humid periods there was every chance that the eastern barrier would occasionally have been permeable. Plumping for the 60 ka exit model was a conservative view stifled by a lack of high-quality dates for scattered suggestions of an Asian AMH presence, such as occurrences of stone tools resembling those of early moderns and even rarer, incomplete and often ambiguous skeletal remains. The ‘modern-looking’ tools that occurred both above and below the 74 ka Toba ash deposit in southern India were disposed of as ‘advanced’ tools of earlier migrants; probably Homo erectus. In retrospect, the established fact of earlier occupation of Eurasia by such ‘primitive’ African migrants, as long ago as ~1.8 Ma in the case of Homo fossils in Georgia, should have encouraged the view that culturally better-endowed AMH would have had less problem in diffusing eastwards once they found an escape route from Africa.
Whatever, the flurry during the last couple of decades of more skeletal and archaeological remains of AMH in Asia, genetic evidence for their interbreeding in the west and east with earlier human groups and, principally, improvements in dating ancient sites suggests a more complex geographic flow. Christopher Bae of the University of Hawaii and colleagues based in the UK, Germany and the US have reviewed this growing wealth of new data to put forward various scenarios for Out of Africa dispersal through Asia (Bae, C.J. et al. 2017. On the origin of modern humans: Asian perspectives. Science, v. 358, p. 1269 (summary); online full paper DOI: 10.1126/science.aai9067). They highlight growing evidence for at least one pre-60 ka dispersal, and probably several, to reach the Levant, Arabia, India, China, Laos, Indonesia, the Philippines and Australia before that date. This tallies with Neanderthal and Denisovan DNA segments within the genomes of living Eurasians that indicate interbreeding before 60 ka.
Bae and colleagues also assemble data that bear on where AMH managed to move out of Africa. They resolve the dispute between routes around the northern shores of the Red Sea and crossing the southern Straits of Bab el Mandab by concluding ‘why not both’. Where the migrations went to is currently suggested by the distribution of sites that reveal either pre- or post-60 ka occupancy. The earlier dispersals may have been dominated by following coastlines along the Mediterranean in North Africa to the Levant and via Bab el Mandab across the Persian Gulf, along the northern Indian Ocean littoral to south-east and east Asia. The later, more ‘adventurous’ movements using both routes led to Europe and deeper into continental Asia and thence to its north east. The review conveniently covers in seven pages much the same geoarchaeological and anthropological ground as Earth-Pages has visited bit-by-bit as it has unfolded since 2000. Clearly, great swathes of Asia have not been explored by palaoanthropologists. As in most geographic sciences there is a tendency to follow up known sites year after year – often decade after decade – to ensure publishable results, and that will consume lots of economic and human resources. It is more risky to try and fill in the gaps, but that basic field work is urgently needed to supply new material.
Vindija cave near Varazdin in Croatia. (credit: Wikipedia)
Work on Neanderthal genomes continues, as expected. The latest news comes from remains of a Croatian female, whose genome has been determined by a team led by members of the Max Planck Institute for Evolutionary Anthropology in Leipzig (Prüfer, K. and 35 others 2017. A high-coverage Neandertal genome from Vindija Cave in Croatia. Science, online; doi:10.1126/science.aao1887). Her full genome is of higher quality than those previously published for Neanderthals. Dated at around 52 ka, her genetics is likely to be closer to those who mated with the ancestors of modern Eurasians. Kay Prüfer and his colleagues suggest that Neanderthals passed on to modern Eurasians genes associated with plasma levels of LDL cholesterol and vitamin D (mainly produced by skin exposure to sunlight, Vitamin D is essential for healthy bones and supports the immune system), together with risk factors for eating disorders, accumulation of visceral fat, rheumatoid arthritis and schizophrenia. Two other interesting possibilities stem from reconsidering genetic data from other Neanderthals, in the light of the new Croatian analysis. Ancestors of an older Neanderthal (122 ka) from the Altai region of Siberia had interbred with genetically modern humans as long ago as 130 ka. Yet the genomes of the Altai and Croatian Neanderthals are surprisingly similar, suggesting that both lived in isolated small groups around three thousand strong (Gibbons, A. 2017. Neanderthal genome reveals greater legacy in the living. Science, v. 358 p. 21).
Also published on-line by Science is a study of the genomes of 7 individual anatomically modern humans from KwaZulu-Natal in South Africa (Schlebusch, C.M. and 11 others 2017. Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago. Science online; doi: 10.1126/science.aao6266). They are three 2000 year-old hunter-gatherers (probably San) and four Iron Age farmers, similar to modern Bantu speakers, from 300 to 500 years ago. Although genomes from modern Khoe-San suggest up to 30% admixture from East Africans and Eurasians, comparison between the two ancient groups suggest a very old divergence among African anatomically modern human (AMH) populations, of the order of 350 to 260 ka ago. This is long before the remains from Ethiopia, widely accepted as the oldest known AMH (190 ka), but roughly the same as recently described fossils from Jebel Irhoud in Morocco (325 to 286 ka) reckoned to be early AMH. The Moroccan humans and now the genetic analysis from much more recent South African skeletons point to a pan-African early evolution of modern people rather than some kind of ‘cradle of humanity’.
Adult male in an Upper Paleolithic (28-30 ka) burial at Sunghir, Russia. (credit: Wikipedia)
More ancient AMH hunter-gatherers (~28 to 30 ka) occur at Sunghir, about 200 km east of Moscow, a settlement that includes several burials – one of which contained a boy and a girl – many with abundant, ornate grave goods. It seems likely that all the interred individuals were related and so an excellent target for DNA analysis. Four individuals with roughly the same 14C age did yield enough for genome sequencing (Sikora, M. and 26 others 2017. Science, online; doi: 10.1126/science.aao1807). It turned out from their mtDNA that none were more closely related than first-cousins or great-grandchildren. The data suggested a relatively small breeding population (~300) that avoided inbreeding and its often negative consequences, possibly through exogamy (a wide mating network) as practiced by living hunter-gatherers. Remarkably, the data also hinted at relationship with earlier (36 ka) individuals from Kostenki about 300 km to the south-west. DNA from several Siberian Neanderthal individuals do suggest that inbreeding had been an issue. Had it been widespread among Neanderthals – risky to infer from such scanty information – that may account for their lack of competitiveness with AMH and eventual demise.
Despite the lingering public image that Neanderthals were not as bright as fully modern humans some had significantly larger brains than we do, albeit with most of the difference being in the rear part of the brain region. So they may have had different powers, such as enhanced vision and awareness of position (proprioception). Because there are few cranial fossils of immature Neanderthals and, for them, little evidence of ages, not much is known about how they developed from birth. A common assumption has been that because their brain was larger post-natal development much have been faster than in modern humans. Set against our slow post-natal development and the faster pace in chimpanzees this assumption has been used in support of limited Neanderthal cognitive abilities.
The El Sidron Neanderthal boy, including a reconstruction of his skull and brain cast. (credit: Antonio Rosas, Museo Nacional de Ciencias Naturales, Madrid, Spain)
The El Sidron cave in Asturias region of northern Spain has yielded fossil remains of a dozen Neanderthals dated at between 49 and 37 ka, the time when anatomically modern humans were also present in Europe. They are among the best studied examples of this human group. Three were of boys, the best preserved of whom is estimated to have died at 7.7 years old from analysis of his dental development (Rosas, A. and 10 others 2017. The growth pattern of Neandertals, reconstructed from a juvenile skeleton from El Sidrón (Spain). Science, v. 357, p. 1282-1287; doi:10.1126/science.aan6463) Analysis of signs of the maturation stage that he had reached, including that of his brain, show no fundamental difference from modern human juveniles in his overall pace of growth. Other workers have found that a similarly aged Homo erectus boy from Kenya had indeed developed more quickly than modern human juveniles.
It’s not much to go on, but the El Sidron boy supports the view that Neanderthals were not much different from us.
You can find more information on migration of modern humans here.
To see traces of where our forebears walked, such as the famous Australopithecusafarensistrackway at Laetoli in Tanzania, the footprints of Neanderthal children in 350 ka old Italian volcanic ash (The first volcanologists? Earth Pages March 2003) or even those of Mesolithic families in estuarine mud is about as heart stopping as it gets for a geologist. But imagine the astonishment of members of a multinational team working on Miocene shore-line sediments on Crete when they came upon a bedding surface covered with what are almost certainly the footprints of another bipedal animal from 5.7 Ma ago (Gierliński, G.D. et al. 2017. Possible hominin footprints from the late Miocene (c. 5.7 Ma) of Crete? Proceedings of the Geologists’ Association, online; https://doi.org/10.1016/j.pgeola.2017.07.006). Trackways preserve a few moments in time, however old they are and the chances of their being preserved are very small, yet they can supply information that is lost from even the best preserved fossil, such as gait, weight, speed and so forth.
Track bearing surface; (b) two footprints in 5.7 Ma old Miocene sediments at Trachilos, Crete (credit: Gierliński, G.D. et al. 2017; Figures 2 and 8)
The tracks clearly indicate that whatever left them was bipedal and lacked claws, and closely resemble those attributed to A. afarensis at Laetoli in a 3.7 Ma old volcanic ash. What they do not resemble closely are those of non-hominin modern primates, such as chimpanzees. They are diminutive compared with adult modern human prints, being about 12.5 cm long (equivalent to a UK child’ shoe size 4 – US size 4.5, EU 20) and about a third to half the size of those at Laetoli. Were they around the age of those at Laetoli or younger there seems little doubt that they would be widely interpreted as being of hominin origin. But being from an island in the Mediterranean as well as far from sites in Africa that have yielded Miocene hominins (Ardipithecus kadabba from Ethiopia, Orrorin from Kenya and Sahelanthropusfrom Chad), such an interpretation is bound to create controversy. Somewhat less controversial might be to regard them as having been created by a late-Miocene primate that convergently evolved a hominin-like upright gait and foot. Being preserved in what seem to be coastal marine sediments, there is probably little chance of body fossils being preserved in the exposed horizon. Since foot bones are so fragile, even if a primate fossil is discovered in the late Miocene of Crete the chances of resolving the issue are pretty remote. Yet fossil primate specialists will undoubtedly beat a well-trodden path to the Trachilos site near Kissamos on Crete
In late July 2017 news emerged that modern humans first reached Australia at least 65 thousand years ago. Confirming that the date of departure from Africa to end up in SE Asia and Australasia was considerable earlier than previously believed, deposits in Sumatra that contain remains of early Home sapiens have yielded even older ages (Westaway, K.E. and 22 others 2017. An early modern human presence in Sumatra 73,000–63,000 years ago. Nature v. 548 online; doi:10.1038/nature23452). This resulted from a re-examination of material from the Padang Caves first excavated more than a century ago by Eugène Dubois, famous for his discovery in Java of the first H. erectus remains. A richly fossiliferous breccia in the Lida Ajer cave yielded a fauna characteristic of a rainforest biome and included two teeth that Dubois considered to be human. Several later palaeontologists confirmed his identification as have hominin specialists in the present Australian-Indonesian-American-British-Dutch-German team. The fossil assemblage has long suggested great antiquity for the site, but only now has it been dated precisely. The dating employed three methods: optically stimulated luminescence dating of quartz grains from the breccia (85±25 to 62±5 ka); uranium-series dating of speleothem including fragments of hollow ‘soda-straw’ stalactites(84±1 to 71±7 ka); uranium-series dating of gibbon and orangutan teeth found together with the human teeth (86±13 to 76±7 ka). Statistical analysis of the age data suggests 73 to 63 ka for the fauna, with a maximum age for deposition of the breccia of 84±1 ka.
Satellite image of Lake Toba in NW Sumatra (at centre), the site of the largest volcanic eruption during the history of human evolution ~71,600 years ago (credit: Wikipedia)
Stone tools which may have been carried by anatomically modern humans into the area have previously been used to suggest a minimum date of the arrival of migrants, though they may have been carried by H. erectus. Remarkably, such tools have been found beneath a thick bed of volcanic ash found throughout southern Asia and in Indian Ocean sediment cores. This has been dated at 71.6 ka and represents the explosive collapse of the caldera now containing Lake Toba in NW Sumatra that was the largest volcanic event in the entire history of the genus Homo. The new age data from Lida Ajer suggests that modern humans were present in its vicinty before the eruption, a view also supported by ‘molecular-clock’ dating of the range of mitochondrial DNA carried by living SE Asian people (79 to 75 ka). So, despite the stupendous magnitude of the Toba eruption is seems likely that some of the migrants survived. Together with the dating of the earliest Australians the Sumatran evidence is at odds with the view, widely held by palaeoanthropologists, that the ‘Out of Africa’ exodus began by crossing the Straits of Bab el Mandab between 74 and 58 ka when global sea-level fell markedly during marine oxygen-isotope Stage 4 (MIS4). A problem with that hypothesis has been that climatic and ecological conditions in southern Asia during MIS4 were unfavourable. But is seems that modern humans were already there and capable of adapting to both the climate shift and to the devastation undoubtedly caused by Toba.
When modern humans first reached Australia has an importance beyond the starting date for the island continent’s archaeology and confirmation that their ancestors are the oldest known migrants from Africa. The first native Australians carried within their genome important information about the minimum date at which some non-Africans interbred with more archaic Neanderthal and Denisovan humans, traces of whose DNA are are present in that of living Australian aborigines. Most dating of when modern humans first reached different parts of the non-African world has relied on the radiocarbon method, which is suspect from beyond 40 to 45 ka as 14C produced earlier has decayed to levels that are now below the practical limit of detection and measurement. It is therefore no accident that the bulk of ‘first-arrival’ dates for Eurasia and Australasia are around 45 ka. In fact, any accurate age, however old, for the earliest skeletal remains only indicates the minimum date of arrival until other remains are discovered.
Reliable dating of earlier events in the Pleistocene relies on other methods, the most important for settings other than speleothem from caves being optically stimulated luminescence (OSL) applied to soil minerals that estimates their time of burial. Briefly, molecules of soil grains made of a mineral such as quartz are ‘charged-up’ with energy by radiation emitted by unstable isotopes in the soil. Exposure to light releases that stored energy in the form of luminescence. Measuring the amount of luminescence emitted by optically stimulated grains therefore gives a measure of the time since they were buried and ceased to be exposed to sunlight.
The Madjedbebe rock shelter in Arnhem Land, Northern Territories, Australia. (Credit: Chris Clarkson, University of Queensland)
A re-evaluation of the Madjedbebe site in the Northern Territory, widely accepted as having yielded Australia’s oldest artefacts in 1989, takes back human occupation more than 20 thousand years before previous estimates (Clarkson and 27 others 2017. Human occupation of northern Australia by 65, 000 years ago. Nature, v. 547, p. 306-310; doi:10.1038/nature22968). The soil profile in the Madjedbebe rock shelter turns out to be littered with artefacts – including hearths, tools and blocks of ochre and reflective mica pigments, plus remnants of plant foods – to a depth of ~2.5 m, with three particularly dense accumulations. Carbon-rich remains are also present throughout the profile which provided a means of accurate calibration and confirmation of OSL dates back as far as the radiocarbon method allows, giving confidence in the older OSL dates that extend to 65.0±5.7 ka in the earliest zone of dense artefact finds. Because the modern DNA of Australia’s first native people shows no sign of mixture with other modern humans, this places the timing of modern human interbreeding with archaic people before this time. The age also predates the range when the continent’s megafauna began to decline to eventual extinction, which supports the view that it was anthropogenic.
See also: Marean, C.W. 2017. Early signs of human presence in Australia. Nature, v. 547, p. 285-287; doi:10.1038/547285a.
How evolution proceeds and species arise are affected by many different processes. But, if members of every generation of the clade that led from the probable common ancestor of ourselves, Neanderthals, Denisovans and other hominins of the last 700 ka or so – widely thought to have been Homo heidelbergensis – were found as perfectly preserved fossils they would show gradually shifting anatomical features that from time to time and place to place would diverge to lead to different species. If, also, every specimen was accurately dated then there would be the last part of the human evolutionary bush laid out in a 3-D graphic. That is never going to be possible, of course. Human fossils are rare and there are few of them that are well-preserved. So the field of human origins throws up surprises on a regular basis, and if palaeoanthropologists were more dogmatic than most of them actually are there would be equally regular, public displays of the eating of hats.
As regards early modern H. sapiens, fossils from a couple of sites in Ethiopia have been the oldest known, at between 160 to 195 ka, for the last 15 years. However, in the 1960s quarry workers at Jebel Irhoud in SW Morocco exposed the infill of a cave network in which were found numerous items of the Levallois stone-tool technology, some human bone fragments that included a brain case and many dismembered and cut bones of prey animals. Initially they were thought to date from about 40 ka and to represent an African form of Neanderthals. Subsequently, re-evaluation of the remains revealed a greater likelihood that they were from modern humans, but too young to be of great interest. An upgraded date of ~160 ka caused them to be considered as peripheral to the core group of Ethiopian early modern humans. DNA analyses then suggested modern humans to have split from Neanderthals about 500 ka ago. Members of the French-Moroccan team that did the original work, accompanied by other scientists, recently re-excavated the site and exhumed a much richer fossil haul that pin-pointed an anatomically modern human (AMH) provenance, albeit with some archaic characteristics (Hublin, J.-J. and 10 others, 2017. New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature, v. 546, p. 289-294; doi:10.1038/nature22336), which can be referred to as ‘pre-modern’ H. sapiens. The bombshell stemming from their work was the precise dating of the fossils and their stratigraphic context by other members of the team (Richter, D. and 11 others. The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Nature, v. 546, p. 293-296; doi:10.1038/nature22335), which yielded 315±34 ka from fire-heated flint fragments and 286±32 ka from a human tooth. Both dates are far older than the previously accepted maximum of 200 ka for AMH.
The early evolution of fully modern humans seems to have spanned the whole of Africa, rather than being set in an Ethiopian heartland, a view partly supported by a fragmentary 260 ka fossil from South Africa bearing close resemblance to the Moroccan individuals. Interestingly, Levallois stone tools, as their name suggests, are widespread in both Africa and Europe at around 300 ka, although that is not proof that AMH migrated out of Africa around 300 ka, for Neanderthals may also have been using a similar flint flaking method (another space to be watched).
See also: Stringer C. & Galway-Witham, J., 2017. On the origin of our species. Nature, v. 546, p. 212-215; doi:10.1038/nature 546212a.
You can find more information on migration of modern humans here.
In September 2015 a barrage of publicity announced the remarkable unearthing of the remains of 15 diminutive hominins, dubbed Homo nadeli, from the floor sediments of an almost inaccessible South African cave, part of the equally hyped ‘Cradle of Humankind’ UNESCO World Heritage Site near Johannesburg. An international team of lithe women speleo-archaeologists was recruited for the excavation, for which the original discoverers were incapably burly. The remains included numerous examples of still articulated intricate bones, such as those of feet and hands, and none show signs of dismemberment by large scavengers. Indeed the discovery chamber was so far from the cave entrance that such animals probably were unaware of their presence. These features and the sheer complexity of the system strongly suggested that cadavers had been deliberately taken to the chamber; implying that the deep penetration had been accomplished using fire-brand illumination. What seized the headlines was the possibility of ritual burial, although sanitary disposal or panicked refuge from predators seem equally, if not more likely.
Lee Burger and the reconstructed skull of Homo naledi
Now yet more fossils have been reported from a separate chamber at a crawling distance about 150 m away from the original but closer to the system’s main entrance (~85 m). These add at least other 3 individuals to the H. nadeli association, with sufficient similarity to indicate that all 18 belong to H. naledi. This wealth of detail enabled the team of authors (Hawks, J. and 37 others 2017. New fossil remains of Homo naledi from the Lesedi Chamber, South Africa. eLife, v. 6, online; http://dx.doi.org/10.7554/eLife.24232) to perform a detailed comparative anatomic analysis of the species. The results are a mosaic, showing some post-cranial affinities with australopithecines, H. habilis, H.floresiensis, H. erectus, Neanderthals and anatomically modern humans, and others, such as the hands and shoulders, that are not well matched with other hominins. Their crania show a similar broad spectrum of resemblances, and as regards dentition they are distinctly primitive. They are also on the small-brained side of the hominin clade. Despite the astonishing abundance of fossil material, not a single artifact was found in the cave system, despite the apparent similarity of its hands to those of ourselves and Neanderthals.
With plenty of scope for speculation, H. nadeli remains enigmatic. The big question looming over the 2015 announcement of the species was its age, the discovers suggesting about 2 Ma, and placing on the direct line of human descent. On the same day as the fossil description there appeared a multi-method dating analysis (Dirks, P.H.G.M. and 19 others 2017. eLife, v. 6, online; http://dx.doi.org/10.7554/eLife.24231.001), which showed that with little doubt that the H. nadeli association was deposited between 236 ka and 335 ka; around the time when anatomically modern humans first emerged and stone tools had undergone a >2 Ma technological evolution. To me, the only sensible conclusion at present is that H. nadeli is another addition to the 6 species living and in some cases coexisting across the late Pleistocene world, and that expansion of ideas beyond that must await DNA analysis; a definite possibility considering the age of the fossils, their seemingly good preservation in a relatively dry cave system and the new possibility of cave soils as well as bones yielding genetic materials. The leader of the research team, Lee Berger of the University of the Witwatersrand now maintains, together with four other members of the research team, that H. nadeli may be a coelacanth-like survivor of Homo’s earliest diversification and that ‘we cannot exclude that this lineage was responsible for the production of Acheulean or Middle Stone Age tool industries’.
Out on the plains countless herbivores fertilise the ground by continual urination and defecation. A friend’s sheep are doing just that in the small field that came with my current home while they are keeping the grass under control. Millions of hectares of prime agricultural land in China are kept fertile through disposal of human night soil from ‘honey wagons’ every day; it is even fed to fishes in small ponds. Such a nice economy also donates the DNA of the animal and plant inhabitants to the soil system. In 2015 analysis of environmental DNA from permafrost in Siberia and Alaska produced ‘bar codes’ for the now vanished ecosystems of what was mammoth steppe during the climate decline to the last glacial maximum and the subsequent warming. The study revealed mammoth and pre-Columbian horse DNA and changes in the steppe vegetation, from which it was concluded that the steppe underwent regional extinction pulses of its megafauna linked to rapid climate ups and downs connected with Dansgaard-Oeschger cycles. It was but a small step to see the potential for studying distribution and timing of various hominins’ occupation of caves from the soils preserved within them, without depending on generally very rare occurrences of human skeletal remains.
Tourists at the entrance to Denisova Cave, Russia (credit: Wikipedia)
The Max Planck Institute for Evolutionary Anthropology in Leipzig, now famous for extracting DNA from Neanderthal, Denisovan and possibly H. antecessor fossils, has applied the environmental DNA approach to sediments from 7 caves in France, Belgium, Spain, Croatia and Russia that span the period from 550 to 14 ka (Slon, V. and 30 others 2017. Neandertal and Denisovan DNA from Pleistocene sediments. Science, v. 356 (online publication); doi:10.1126/science.aam9695). The sites had previously yielded fossils and/or artefacts. All of them contained mitochondrial DNA from diverse large mammals, four including archaic human genetic material supplied by Neanderthal individuals and Denisovans in the case of the Denisova cave. A key finding was Neanderthal mtDNA in one sedimentary layer that contained no skeletal remains – decay of a body was probably not involved. In two cases the DNA was from more than one individual. A variety of tests showed that surprisingly large quantities of DNA survive in soil and that it is spread evenly in sediment rather than being present in spots – an indication of derivation from urine, excreta or decayed soft tissue.
Although the study does not add to knowledge of hominin genetics, it confirms that the methodology is sufficiently advanced and efficient to detect hominin presence in fossil-free sediment. So this approach seems set to become a standard for many sites, such as that from California reported in the previous post, which suggest a human influence, or any cave sediments for that matter. Although skeletal remains are essential for reconstruction of bodily characteristics, hominin phylogeny seems set to cut loose from fossils. Hitherto suspected species’ presence in the time period where DNA analysis is feasible may be detected, such as Asian H. erectus. It may become possible to map or extend the geographic ranges of Denisovans and Neanderthals. Perhaps species new to science will emerge.
In 1991-2 palaeontologists excavated a site near San Diego, California where broken bones had been found. These turned out to be the disarticulated remains of an extinct mastodon. One feature of the site was the association of several large cobbles with bones of large limbs that seemed to have been smashed either to extract marrow or as source of tool-making material. The cobbles showed clear signs or pounding, such as loss of flakes – one flake could be fitted exactly to a scar in a cobble – pitted surfaces and small radiating fractures. The damage to one cobble suggested that it had been used as an anvil, the others being hammer stones. Broken pieces of rock identical to the hammer stones were found among the heap of bones. No other artefacts were found, and the bones show no sign of marks left by cutting meat from them with stone tools. The breakage patterns of the bones included spiral fractures that experimental hammering of large elephant and cow bones suggest form when bone is fresh. Other clear signs of deliberate breakage are impact notches and small bone flakes. Two detached, almost spherical heads of mastodon femora suggest that marrow was the target for the hammering and confirmed the breakage was deliberate.
Artist’s impression of American mastodon. (credit: Wikipedia)
Since the sediment stratum in which the remains occurred consists of fine sands and silt, typical of a low-energy river system, the chances that the cobbles had been washed into association with the mastodon are very small. The interpretation of the site is that it was the result of opportunistic exploitation of a partial carcase of a young adult mastodon by humans. In the early 1990s attempts were made to date the bones using the radiocarbon method, but failed due to insufficient preserved collagen. That the site may have been much older than the period of known occupation of North America by ancestors of native people (post 14.5 ka) emerged from attempts at optically stimulated luminescence dating of sand grains that can suggest the age of burial. These suggested burial by at least 60 to 70 ka ago. It was only when the uranium-series disequilibrium method was used on bone fragments that full significance of the site emerged. The results indicated that they had been buried at 130.7±9.4 ka (Holen, S.R. and 10 others 2017. A 130,000-year-old archaeological site in southern California, USA. Nature, v. 544, p. 479—493; doi:10.1038/nature22065 – full paper and supplements available free)
Not only is the date almost ten times that of the earliest widely accepted signs of Homo sapiensin the Americas, the earliest anatomically modern humans known to have left Africa are around the same age, but restricted to the Levant. The earliest evidence that modern humans had reached East Asia and Australasia through their eastward migration out of Africa is no more than 60 ka. The date from southern California is around the start of the interglacial (Eemian) before the one in which we live now. It may well have been possible then, as ~14 ka ago, to walk across the Bering Straits due to low sea level, or even by using coast-hugging boats – hominins had reached islands in the Mediterranean and the Indonesian peninsula certainly by 100 ka, and probably earlier. But whoever exploited the Californian mastodon marrow must have been cold-adapted to achieve such a migration. While the authors speculate about ‘archaic’ H. sapiens the best candidates would have been hominins known to have been present in East Asia: H. erectus, Neaderthals and the elusive Denisovans.
Surely there will be reluctance to accept such a suggestion without further evidence, such as tools and, of course, hominin skeletal remains. But these long-delayed findings seem destined to open up a new horizon for American palaeoanthropology, at least in California.
You can find more information on hominin migration here.
For many years anthropologists were certain that the Americas remained outside the human realm until the great icecap of North America had begun to melt decisively. This view stemmed partly from the only conceived route being across the exposed floor of the Bering Sea when sea-level had fallen to leave it as a landmass known as Beringia. The other literal stumbling block had been the glacial blockage of the only lowland corridor from Alaska to the Great Plains which roughly follows the Alberta – British Columbia border in Canada. There is abundant evidence that the corridor did not become ice-free until about 13 ka, an important fact that for a long while bolstered the Clovis-First hypothesis, from the eponymous and highly distinctive stone tools that date back to just after that time. After a long, sturdy rearguard action by its devotees that view was transcended by finds of earlier tools with dates as old as 15.5 ka that extend close to the southernmost tip of South America. Studies of Y-chromosome DNA from living First Nations men that suggested that all early Americans stemmed from 4 separate colonising populations who may have entered via Beringia by different routes, including along the Pacific coast. A possible common ancestor of all native Americans has turned up from the mitochondrial and Y-chromosome DNA of a fossil skeleton from near Lake Baikal in Siberia who lived about 24 ka ago. But yet another twist has emerged from the Yukon Territory of Northern Canada.
Beringia Land Bridge. Animation of its development from 21.000 BC to modern times.(Photo credit: Wikipedia)
Since 1987 it has been known that animal bones with clear signs of butchery occurred in the Bluefish Cave on the Yukon – Alaska border. Dating of the bones by the 14C method seemed to support human occupation there during the Last Glacial Maximum; highly controversial at the time, in the absence of any other sites of that age in the whole Americas. The material has now been re-examined and dated by a more advanced radiocarbon method (Bourgeon, L. et al. 2017. Earliest human presence in North America dated to the Last Glacial Maximum: new radiocarbon dates from Bluefish Caves, Canada. PLoS ONE, v. 12; doi:10.1371/journal.pone.0169486). This work has confirmed the earlier view since the ages of bones range from 24 to 12 ka. But the discovery of what seems long-term occupation under the most arduous glacial conditions is not the only outcome of the research. One hypothesis for the genetic diversity among living indigenous people of the Americas is that their forebears, the first people of the Americas, may have been from genetically isolated populations stranded on Beringia, yet surviving eventually to migrate southward once climate warmed. The ‘Beringian standstill hypothesis’ suggest that the small population underwent genetic drift for about eight thousand years, their descendants inheriting the genetic diversity produced by this process. Bluefish Cave is probably where some of those pioneers waited-out the Ice Age
Teeth are the most likely parts of skeletons to survive for long periods because of their armour by a layer of enamel made of hydroxyapatite (Ca5(PO4)3(OH)). Dental enamel is the hardest material in the bodies of vertebrate animals and lies midway between fluorite and feldspar on Moh’s scale of hardness (value 5). Like the mineral apatite, teeth survive abrasion, comminution and dissolution for long periods in the surface environment. Subdivision of fossil hominin species and even among different groups of living humans relies to a marked extent on the morphology of their teeth’s biting and chewing surface. Although there are intriguing examples in Neolithic jawbones of dental cavities having been filled it is rather lack of attention to teeth that characterises hominin fossils. As well as horrifying signs of mandibular erosion due to massive root abscesses, a great many hominin remains carry large accumulations of dental plaque or calculus made of mineralised biofilm laid down by oral bacteria. Even assiduous brushing only delays the build up. Grisly as this inevitability might seem, plaque is an excellent means of preserving not only the bacteria but traces of what an individual ate. As fossil DNA is a guide to ancestry and relatedness among fossil hominins, so far going back to about 430 ka in the case of a Spanish Homo heidelburgensis, plaque potentially may reveal details of diet and to some extent social behaviour elaborating beyond the possibilities presented by carbon isotopes and dental wear patterns.
Plaque deposits have already shown that Neanderthals had a very varied vegetable diet and that they cooked their food, the sugars thereby released encouraging bacterial biofilms. There have even been hints that they used medicinal herbs, such as yarrow and chamomile. Now a large multinational team of scientists has taken this fascinating line of study a step further using short DNA fragments to identify the actual oral microbes and even plant and animal species that dominated the diets of 8 cave-dwelling Neanderthals found in Spain, Belgium and Italy (Weyrich, L.S. and 30 others 2017. Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus. Nature, v. 543; online doi:10.1038/nature21674). The Spanish individuals found in El Sidrón cave seem to have been mainly vegetarian (mushrooms, pine nuts and edible moss) whereas two from the Spy cave in Belgium feasted on wooly rhinoceros and mouflon sheep. One of the El Sidrón Neanderthals had a dental abscess, and was probably in great pain, and whose calculus contained evidence of ingestion of tissue from poplar trees, known to contain salicylic acid (the active ingredient in aspirin): an example of self-medication. The unfortunate individual was also suffering from acute diarrhoea brought on by a eukaryote parasite (microsporidium). Astonishingly, DNA from several plant fungi, including Penicillium rubens (penicillin) also occurred in this individual’s calculus, from eating mouldy plant material: predating modern antibiotics by more than forty-five thousand years!
More predictable findings from the unfortunate El Sidrón individual was a spectrum of common plaque colonising bacteria. But another surprise was Methanobrevibacter oralis, an archaea common in the human mouth ecosystem, for which a complete genome was reconstructed. It is different from that in the Methanobrevibacter oralis found in living humans and the team were able to use a molecular clock approach to date the divergence between the two sub-species. This seems to have occurred between 112-143 ka ago, long after the divergence of Neanerthals and anatomically modern humans, judged to be around 450 to 750 ka ago. The authors suggest that ‘commensal microbial species were transferred between the two hosts during subsequent interactions, potentially in the Near East’. Two alternative ‘interactions’ occurred to one commentator: kissing or exchange of chewed food (Callaway, E, 2017. Plaque DNA hints at Neanderthal lifestyle. Nature, v. 543, p. 163). Intriguingly the date, albeit imprecise, overlaps with estimates for the timing of Neanderthal – modern human interbreeding as the latter began to leave Africa: not only do living non-Africans share genes with Neanderthals, the may also share saliva and oral bacteria.
For more information on recent human evolution see here.









")