
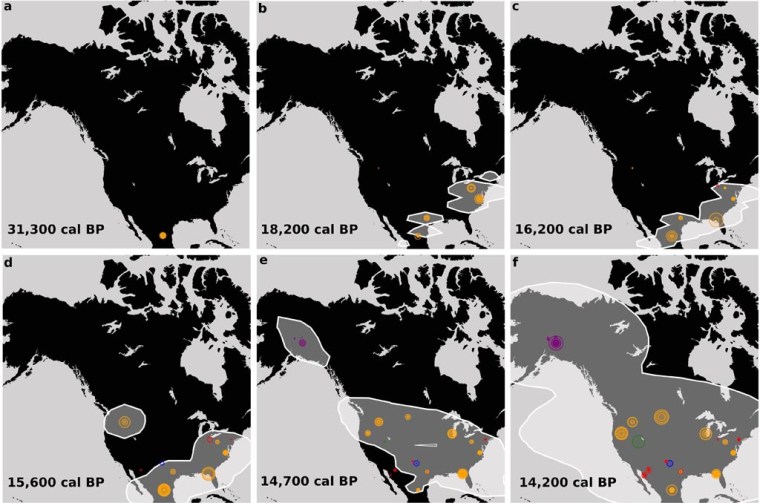
When and how humans first migrated into the Americas are issues that have exercised anthropologists for the last two decades, often sparking off acrimonious debate. In the 20th century both seemed to well established: hunters using the celebrated Clovis fluted spear blades arrived first, no earlier than 13 ka ago. The Beringia land bridge across what is now the Bering Strait was exposed by falling sea level as early as 30 thousand years ago in the lead-up to the last glacial maximum (LGM) to link eastern Siberia and Alaska. However, ice sheets expanding to the south-west of the main area of glaciation on the Canadian Shield barred passage through Interior Alaska and NW Canada. Only around 13 ka had a N-S ice-free corridor opened through the mountains during glacial retreat. Nevertheless, humans had entered Alaska at least ten thousand years earlier, during the LGM, to occupy caves in its western extremity: Alaska was habitable but they were stuck there.
In the early 21st century, it became clear that the ‘Clovis First’ hypothesis was mistaken. Sediments in Texas that contained Clovis blades were found to be underlain by those of an older culture, reliably dated to about 15.5 ka. Furthermore, analysis of the DNA of all groups of native Americans (north and south) indicated a last common ancestor in Siberia more than 30 ka ago: they descended from that ancestor outside of Asia. More recently excavated sites in Mexico and Chile point to human populations having reached there as early at 33 ka (see: Earliest Americans, and plenty of them; July 2020), and there is a host of pre-Clovis sites in North and Central America dating back to 18.2 ka. Such ancient groups could not have walked from the Beringia land bridge because the present topographic grain in the Western Cordillera would have been blocked by ice since about 25 thousand years ago. The only viable possibility was that they followed the Alaskan coast to move southwards, either in boats or over sea ice.
A new focus on when such journeys would have been feasible was published in February 2023 (Praetorius, S.K et al. 2023. Ice and ocean constraints on early human migrations into North America along the Pacific coast. Proceedings of the National Academy of Science, v. 120, article e2208738120; DOI: 10.1073/pnas.2208738120). One advantage of moving along the coast is that, though it would be pretty cold, the warming effect of the Pacific Ocean would make it more bearable than travelling inland, where winter temperatures even today regularly reach -50°C. More important, there would be no shortage of food; fish, marine mammals and shellfish abound at the ice margin or onshore, at any season. But a coastal route may not have been possible at all times during the period either side of the LGM. Large glaciers still reach the ocean from Alaska and there is little more perilous than crossing the huge crevasse fields that they present. Boating would have been highly dangerous because of continual calving of icebergs from extensive ice shelves. Moreover, the Alaska Coastal Current flows northwards and would likely have sped up during episodes of glacial melting as the current is affected by fresh water influx. Yet there may sometimes have been episodes of open water at the ice front frozen to relatively flat sea ice in winter. That would making boat- or foot travel relatively safe. Sea ice would also make glacier-free islands accessible for encampments over the harsh winters or even for hundreds of years, with plenty of marine food resources.
Summer Praetorius of the US Geological Survey and colleagues from Woods Hole Oceanographic Institution, Oregon State University, and the Universities of California (Santa Cruz) and Oregon have attempted to model conditions since 32.5 ka ago in coastal waters off Northwest America. They used simulations of the behaviour of the Alaska Coastal Current during varying climate conditions before and during the LGM, while glaciers were in retreat that followed and during the Holocene. Their modelling is based on the effects of changing sea level and water salinity on general circulation in the Northern Pacific. The relative abundance of sea ice can be tracked using variation in an alkenone produced by phytoplankton that wax and wane according to sea-surface temperature and sea-ice cover. The other input is the well-documented changing extent of continental glaciation in Alaska and the Yukon Territory. Based on their model they estimate that the most favourable environmental conditions for coastal migration occurred just before the LGM (24.5 to 22 ka) and between 16.4 and 14.8 ka during the initial stages of warming and extensive melting of ice sheets. The Alaskan Coastal Current probably doubled in intensity during the LGM making the use of boats highly dangerous
By 35 ka ocean-going boats are known to have been used by people in northern Japan. Traversing sea-ice was the way in which Inuit people occupied all the Arctic coastal areas of North America and Greenland during the last five thousand years, and is the form of travel favoured by the authors. It is not yet possible to prove and date such coastal journeys because campsites or settlements along the coast would now be inundated by 100 m of post-glacial sea-level rise. Yet such migration was necessary to establish settlements at lower latitudes in North America and Mexico in the period when overland routes from Beringia were blocked by ice sheets. By 32.5 ka falling sea level probably made it possible to cross the Bering Strait for the first time and for the next 7.5 ka an ice-free corridor made it possible for the rest of North America and points further south to be reached on-foot from Alaska. That window of opportunity might have allowed humans to have reached Mexico and South America, where the earliest dates of occupation have been found. But many of the early sites across North America date to the period (25 to 13 ka) when overland access was blocked. Of course, those sites might have been established by expansion from the very earliest migrants who crossed the Beringia land bridge and took advantage of overland passage before 25 ka. But if later migrants from Asia could follow the coastal route, then it seems likely that they did. Later Inuit spread along the shores of the Arctic Ocean since 5000 years ago probably with a material culture little different from that of the earlier migrants from Siberia.


















{kind=link}