Hominins have been making and using stone tools since at least 3.4 Ma, as shown by cut marks on bones and stone artefacts themselves. I use the sack term ‘hominin’ because the likely makers and users of the oldest tools are either australopithecines or paranthropoids, there being no fossils designates to the genus Homo of late-Pliocene age. So it might seem un-newsworthy to report that the oldest tools deliberately made from bone are now known to occur in 1.5 Ma old sediments from the famous sedimentary sequence at Olduvai Gorge in Tanzania (de la Torre, I and 8 others2025. Systematic bone tool production at 1.5 million years ago. Nature, v. 639; DOI: 10.1038/s41586-025-08652-5). To be clear, there is abundant evidence that hominins had used bones, especially sturdy long bones, for digging perhaps, much earlier in hominin history. Again, paranthropoids have been implicated in their use. The bones found at Olduvai actually show signs of manufacture into useful objects prior to their use: they show clear signs of knapping to produce points and blades. The bones are among the sturdiest known from the Pleistocene, being from elephants and hippos. Before de la Torre and colleagues found what is essentially a bone-tool factory, it was thought that systematic use of bones in such a sophisticated manner only arose between 400 to 250 ka ago among early Homo in Europe. Sadly, fossils of whoever made the tools were not found at the site. Once again, paranthropoids as well as early Homo are known to have cohabited the area at that time.
‘Front, back and side’ views of a 1.5 Ma old tool made from an elephant humerus – its upper foreleg. The scale bar represents 5 cm. (Credit: de la Torre et al.; Fig 3a)
Bifacial Acheulean stone artefacts first appear in the rock record about 300 ka before these bone tools were made. So one idea that the authors put forward is that the same kind of stone knapping technique was transferred to the more abundant massive bones of the East African Pleistocene megafauna (in the absence or rarity of suitable blocks of stone?). But it remains unclear whether or not such tools were simply selected from very large bones smashed to get at their nutritious marrow. The first possibility implies a cultural shift, whereas the latter points simply to expedience. The authors are at pains to point out that the curious million-year gap in the record of bone tools may be ascribed either to the disappearance of bone technology or simply to archaeologists who worked elsewhere having not regarding bone fragments as the products of skills. That applies equally to earlier times, when bones were indeed used, though with not so much in the way of a ‘mental template’. As de la Torre et al. conclude ‘Future research needs to investigate whether similar bone tools were already produced in earlier times, persisted during the Acheulean and eventually evolved into Middle Pleistocene bone bifaces similar in shape, size and technology to their stone counterparts’.
Evidence from Dmanisi in Georgia that Homo erectus may have been the first advanced hominin to leave Africa about 1.8 Ma ago was a big surprise (see: First out of Africa?November 2003). Remains of five individuals included one skull of an aged person who face was so deformed that he or she must have been cared for by others for many years. So, a second surprise from Dmanisi was that human empathy arose far earlier than most people believed. Since 2002 there has been only a single further find of hominin bones of such antiquity, at Longgudong in central China. For the period between 1.0 and 2.0 Ma eight other sites in Eurasia have yielded hominin remains. If finds of stone tools and evidence of deliberate butchery – cut marks on prey animals’ bones – are accepted as tell-tale signs, the Eurasian hominin record is considerably larger, and longer,. There are 11 Eurasian sites that have yielded such evidence – but no hominin remains – that are older than Longgudong: in Russia, China, the Middle East, North Africa and northern India. The oldest, at Masol in northern India is 2.6 Ma old. In January 2025 the earliest European evidence for hominin activity was reported from Grăunceanu in Romania (Curran, S.C. and 15 others 2025. Hominin presence in Eurasia by at least 1.95 million years ago. Nature Communications, v. 16, article 836; DOI: 10.1038/s41467-025-56154-9) in the form of animal bones showing clear signs of butchery, as well as stone tools, but no hominin fossils.
Animal bones showing cut marks from the 1.95 Ma old Grăunceanu site in Romania. (Credit: Curran et al. 2025, Figs 2A and C)
There were stone-tool makers who butchered prey in Africa as early as 3.4 Ma ago (see: Stone tools go even further back; May 2015), but without direct evidence of which hominin was involved. Several possible candidates have been suggested: Australopithecus; Kenyanthropus; Paranthropus. The earliest known African remains of H. erectus have been dated at around 2.0 Ma. So, all that can be said with some certainty about the pre-2 Ma migrants to Eurasia, until fossils of that antiquity are found, is that they were hominins of some kind: maybe advanced australopithecines, paranthropoids or early humans. Those from Longgudong and Dmanisi probably are early Homo erectus, and 2 others (1.7 and 1.6 Ma) from China have been designated similarly. Younger, pre-1.0 Ma Eurasian hominins from Israel, Indonesia, Spain and Turkey are currently un-named at the species level, but are allegedly members of the genus Homo.
So, what can be teased from the early Eurasian hominin finds? Some certainly travelled thousands of kilometres from their assumed origins in Africa, but none penetrated further north than about 50°N. Perhaps they could not cope with winters at higher latitudes, especially during ice ages. To reach as far as eastern and western Eurasia suggests that dispersal following exit from Africa would have taken many generations. There is no reason to suppose continual travel; rather the reverse, staying put in areas with abundant resources while they remained available, and then moving on when they became scarce. Climate cycles, first paced at around 40 ka (early Pleistocene) then at around 100 ka (mid Pleistocene and later), would have been the main drivers for hominin population movements, as it would have been for game and vegetation.
After about 3 Ma the 40 ka climate cyclicity evolved to greater differences in global temperature between glacial and interglacial episodes, and even more so after the mid Pleistocene transition to 100 ka cycles (see Wikipedia entry for the mid-Pleistocene Transition). Thus, it seems likely that chances of survival of dispersed bands of hominins decreased over hundreds of millennia. Could populations have survived in particularly favourable areas; i.e. those at low latitudes? If so did both culture and the hominins themselves evolve? Alternatively, was migration in a series of pulses out of Africa and then dispersal in all directions, most ending in regional extinction? Almost certainly, pressures to leave Africa would have been driven by climate, for instance by increased aridity as global temperatures waned and sea-level falls made travel to Eurasia easier. There may also have been secondary, shorter migrations within Eurasia, again driven by environmental changes. Without more data from newly discovered sites we can go little further. Within the 35 known, pre-1 Ma hominin sites there are two clusters: southern and central China, and the Levant, Turkey and Georgia. Could they yield more developments? A 2016 article in Scientific American about Chinese H. erectus finds makes particularly interesting reading in this regard.
Neanderthals and the elusive Denisovans began to establish permanent Eurasian ranges, after roughly 600 ka ago. Both groups survived until after first contact with waves of anatomically modern humans in the last 100 ka, with whom some interbred before vanishing from the record. However, evidence from the DNA of both groups suggests an interesting possibility. Before the two groups split genetically, their common ancestors (H. heidelbergensis or H. antecessor?) apparently interbred with genetically more ancient Eurasian hominins (see Wikipedia entry for Neanderthal evolution). This intriguing hint suggests that more may be discovered when substantial remains of Denisovans – i.e. more than a few teeth and small bones – are discovered and yield more DNA. My guess is such a future development will stem from analysis of early hominin remains in China, currently regarded as H. erectus. See China discovers landmark human evolution fossils. Xinhua News Agency 9 December 2024)
A fully revised edition of Steve Drury’s book Stepping Stones: The Making of Our Home World can now be downloaded as a free eBook
Refugees from the Middle East migrating through Slovenia in 2015. Credit: Britannica
During the Pliocene (5.3 to 2.7 Ma) there evolved a network of various hominins, with their remains scattered across both the northern and southern parts of that continent. The earliest, though somewhat disputed hominin fossil Sahelanthropus tchadensis hails from northern Chad and lived around 7 Ma ago, during the late Miocene, as did a similarly disputed creature from Kenya Orrorin tugenensis (~5.8 Ma). The two were geographically separated by 1500 km, what is now the Sahara desert and the East African Rift System. The suggestion from mtDNA evidence that humans and chimpanzees had a common ancestor, the uncertainty about when it lived (between 13 to 5 Ma) and what it may have looked like, let alone where it lived, makes the notion debateable. There is even a possibility that the common ancestor of humans and the other anthropoid apes may have been European. Its descendants could well have crossed to North Africa when the Mediterranean Sea had been evaporated away to form the thick salt deposits that now lie beneath it: what could be termed the ‘Into Africa’ hypothesis. The better known Pliocene hominins were also widely distributed in the east and south of the African continent. Wandering around was clearly a hominin predilection from their outset. The same can be said about humans in the general sense (genus Homo) during the Early Pleistocene when some of them left Africa for Eurasia. Artifacts dated at 2.1 Ma have been found on the Loess Plateau of western China, and Georgia hosts the earliest human remains known from Eurasia. Since them H. antecessor, heidelbergensis, Neanderthals and Denisovans roamed Eurasia. Then, after about 130 ka, anatomically modern humans progressively populated all continents, except Antarctica, to their geographic extremities and from sea level to 4 km above it.
There is a popular view that curiosity and exploration are endemic and perhaps unique to the human line: ‘It’s in our genes’. But even plants migrate, as do all animal species. So it is best to be wary of a kind of hominin exceptionalism or superior motive force. Before settled agriculture, simply diffusion of populations in search of sustenance could have achieved the enormous migrations undertaken by all hominins: biological resources move and hunter gatherers follow them. The first migration of Homo erectus from Africa to northern China by way of Georgia seems to taken 200 ka at most and covered about ten thousand kilometres: on average a speed of only 50 m per year! That achievement and many others before and later were interwoven with the evolution of brain size, cognitive ability, means of communication and culture. But what were the ultimate drivers? Two recent papers in the journal Nature Communications make empirically-based cases for natural forces driving the movement of people and changes in demography.
The first considers hominin dispersal in the Palaearctic biogeographic realm: the largest of eight originally proposed by Alfred Russel Wallace in the late 19th century that encompasses the whole of Eurasia and North Africa (Zan, J. et al. 2024. Mid-Pleistocene aridity and landscape shifts promoted Palearctic hominin dispersals. Nature Communications, v. 15, article 10279; DOI: 10.1038/s41467-024-54767-0). The Palearctic comprises a wide range of ecosystems: arid to wet, tropical to arctic. After 2 Ma ago, hominins moved to all its parts several times. The approach followed by Zan et al. is to assess the 3.6 Ma record of the thick deposits of dust carried by the perpetual westerly winds that cross Central Asia. This gave rise to the huge (635,000 km2) Loess Plateau. At least 17 separate soil layers in the loess have yielded artefacts during the last 2.1 Ma. The authors radiocarbon dated the successive layers of loess in Tajikistan (286 samples) and the Tarim Basin (244 samples) as precisely as possible, achieving time resolutions of 5 to 10 ka and 10 to 20 ka respectively. To judge variations in climate in these area they also measured the carbon isotopic proportions in organic materials preserved within the layers. Another climate-linked metric that Zan et al. is a time series showing the development of river terraces across Eurasia derived from the earlier work of many geomorphologists. The results from those studies are linked to variations through time in the numbers of archaeological sites across Eurasia that have yielded hominin fossils, stone tools and signs of tool manufacture, many of which have been dated accurately.
The authors use sophisticated statistics to find correlations between times of climatic change and the signs of hominin occupation. Episodes of desertification in Palaearctic Eurasia clearly hindered hominins’ spreading across the continent either from west to east of vice versa. But there were distinct, periodic windows of climatic opportunity for that to happen that coincide with interglacial episodes, whose frequency changed at the Mid Pleistocene Transition (MPT) from about 41 ka to roughly every 100 ka. That was suggested in 2021 to have arisen from an increased roughness of the rock surface over which the great ice sheets of the Northern Hemisphere moved. This suppressed the pace of ice movement so that the 41 ka changes in the tilt of the Earth’s rotational axis could no longer drive climate change during the later Pleistocene, despite the fact that the same astronomical influence continued. The succeeding ~100 ka pulsation may or may not have been paced by the very much weaker influence of Earth changing orbital eccentricity. Whichever, after the MPT climate changes became much more extreme, making human dispersal in the Palearctic realm more problematic. Rather than hominin’s evolution driving them to a ‘Manifest Destiny’ of dominating the world vastly larger and wider inorganic forces corralled and released them so that, eventually, they did.
Much the same conclusion, it seems to me, emerges from a second study that covers the period since ~ 9 ka ago when anatomically modern humans transitioned from a globally dominant hunter-gatherer culture to one of ‘managing’ and dominating ecosystems, physical resources and ultimately the planet itself. (Wirtz, K.W et al. 2024. Multicentennial cycles in continental demography synchronous with solar activity and climate stability. Nature Communications, v. 15, article 10248; DOI: 10.1038/s41467-024-54474-w). Like Zan et al., Kai Wirtz and colleagues from Germany, Ukraine and Ireland base their findings on a vast accumulated number (~180,000) of radiocarbon dates from Holocene archaeological sites from all inhabited continents. The greatest number (>90,000) are from Europe. The authors applied statistical methods to judge human population variations since 11.7 ka in each continental area. Known sites are probably significantly outweighed by signs of human presence that remain hidden, and the diligence of surveys varies from country to country and continent to continent: Britain, the Netherlands and Southern Scandinavia are by far the best surveyed. Given those caveats, clearly this approach gives only a blurred estimate of population dynamics during the Holocene. Nonetheless the data are very interesting.
The changes in population growth rates show distinct cyclicity during the Holocene, which Wirtz et al. suggest are signs of booms and busts in population on all six continents. Matching these records against a large number of climatic time series reveals a correlation. Their chosen metric is variation in solar irradiance: the power per unit area received from the Sun. That has been directly monitored only over a couple of centuries. But ice cores and tree rings contain proxies for solar irradiance in the proportions of the radioactive isotopes 10Be and 14C contained in them respectively. Both are produced by the solar wind of high-energy charged particles (electrons, protons and helium nuclei or alpha particles) penetrating the upper atmosphere. The two isotopes have half-lives long enough for them to remain undecayed and thus detectable for tens of thousand years. Both ice cores and tree rings have decadal to annual time resolutions. Wirtz et al. find that their crude estimates of booms and busts in human populations during the Holocene seem closely to match variations in solar activity measured in this way. Climate stability favours successful subsistence and thus growth in populations. Variable climatic conditions seem to induce subsistence failures and increase mortality, probably through malnutrition.
A nice dialectic clearly emerges from these studies. ‘Boom and bust’ as regards populations in millennial and centennial to decadal terms stem from climate variations. Such cyclical change thus repeatedly hones natural selection among the survivors, both genetically and culturally, increasing their general fitness to their surroundings. Karl Marx and Friedrich Engels would have devoured these data avidly had they emerged in the 19th century. I’m sure they would have suggested from the evidence that something could go badly wrong – negation of negation, if readers care to explore that dialectical law further . . . And indeed that is happening. Humans made ecologically very fit indeed in surviving natural pressures are now stoking up a major climatic hiccup, or rather the culture and institutions that humans have evolved are doing that.
For the last forty thousand years anatomically modern humans have been the only primates living on Planet Earth with a sophisticated culture; i.e. using tools, fire, language, art etcetera. Since Homo sapiens emerged some 300 ka ago, they joined at least two other groups of humans – Neanderthals and Denisovans – and not only shared Eurasia with them, but interbred as well. In fact no hominin group has been truly alone since Pliocene times, which began 5.3 Ma ago. Sometimes up to half a dozen species occupied the habitable areas of Africa. Yet we can never be sure whether or not they bumped into one another. Dates for fossils are generally imprecise; give or take a few thousand years. The evidence is merely that sedimentary strata of roughly the same age in various places have yielded fossils of several hominins, but that co-occupation has never been proved in a single stratum in the same place: until now.
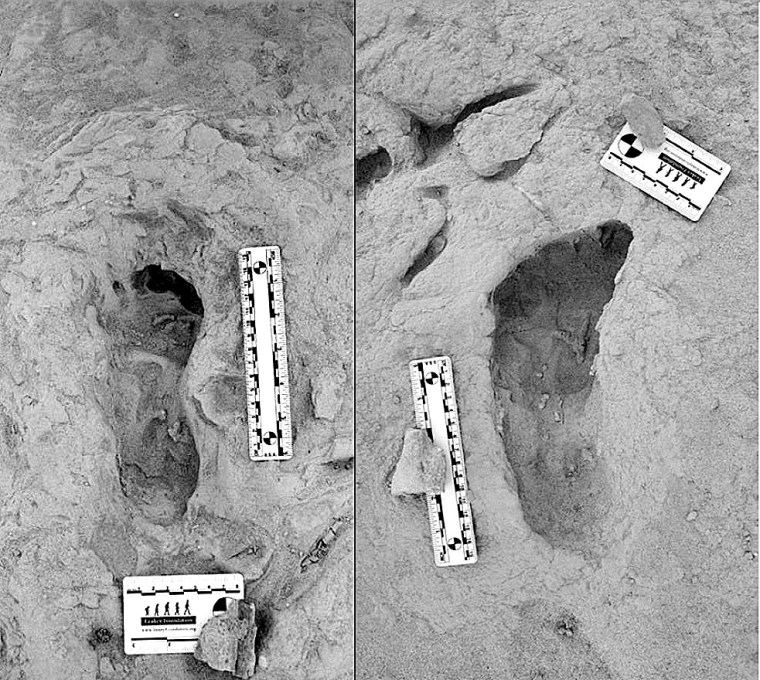
Footprints from Koobi Fora: left – right foot of H. erectus; right – left foot of Paranthropus boisei. Credit: Kevin Hatala. Chatham University
The Koobi Fora area near modern Lake Turkana has been an important, go-to site, courtesy of the Leakey palaeoanthropology dynasty (Louis and Mary, their son and daughter-in-law Richard and Meave, and granddaughter Louise). They discovered five hominin species there dating from 4.2 to 1.4 Ma. So there was a chance that this rich area might prove that two of the species were close neighbours in both space and time. In 2021 Kenyan members of the Turkana Basin Institute based in Nairobi spotted a trackway of human footprints on a bedding surface of sediments that had been deposited about 1.5 Ma ago. Reminiscent of the famous, 2 million years older Laetoli trackway of Australopithecus afarensis in Tanzania, that at Koobi Fora is scientifically just as exciting for it shows footprints of two hominin species Homo erectus and Paranthropus boisei who had walked through wet mud a few centimetres below the surface of Lake Turkana’s ancient predecessor (Hatala, K.G. and 13 others, 2024. Footprint evidence for locomotive diversity and shared habitats among early Pleistocene hominins. Science, v. 386, p. 1004-1010; DOI: 10.1126/science.ado5275). The trackway is littered with the footprints of large birds and contains evidence of zebra.
One set of prints attributed to H. erectus suggest the heels struck the surface first, then the feet rolled forwards before pushing off with the soles: little different from our own, unshod footprints in mud. They are attributed to H. erectus. The others also show a bipedal gait, but different locomotion. The feet that made them were significantly flatter than ours and had a big toe angled away from the smaller toes. They are so different that no close human relative could have made them. The local fossil record includes paranthropoids (Paranthropus boisei), whose fossil foot bones suggest an individual of that speciesmade those prints. It also turns out that a similar, dual walkers’ pattern was found 40 km away in lake sediments of roughly the same age. The two species cohabited the same terrain for a substantial period of time. As regards the Koobi Fora trackway, it seems the two hominins plodded through the mud only a few hours apart at most: they were neighbours.
Artists’ reconstructions of: left – H. erectus; right – Paranthropus boisei. Credits: Yale University, Roman Yevseyev respectively
From their respective anatomies they were very different. Homo erectus was, apart from having massive brow ridges, similar to us. Paranthropus boisei had huge jaws and facial muscles attached to a bony skull crest. So how did they get along? The first was probably omnivorous and actively hunted or scavenged meaty prey: a bifacial axe-wielding hunter-gatherer. Paranthropoids seem to have sought and eaten only vegetable victuals, and some sites preserve bone digging sticks. They were not in competition for foodstuffs and there was no reason for mutual intolerance. Yet they were physically so different that intimate social relations were pretty unlikely. Also their brain sizes were very different, that of P. Boisei’s being far smaller than that of H. erectus , which may not have encouraged intellectual discourse. Both persist in the fossil record for a million years or more. Modern humans, Neanderthals and Denisovans, as we know, sometimes got along swimmingly, possibly because they were cognitively very similar and not so different physically.
Since many hominin fossils are associated with riverine and lake-side environments, it is surprising that more trackways than those of Laetoli and Koobi Fora have been found. Perhaps that is because palaeoanthropologists are generally bent on finding bones and tools! Yet trackways show in a very graphic way how animals behave and interrelate with their environment, for example dinosaurs. Now anthropologists have learned how to spot footprint trace fossils that will change, and enrich the human story.
Charles Darwin famously suggested that humans evolved from apes, and since great apes (chimpanzees, bonobos and gorillas) live in Africa he reckoned it was probably there that the human ‘line’ began. Indeed, the mitochondrial DNA of chimpanzees (Pan troglodytes) is the closest to that of living humans. Palaeoanthropology in Africa has established evolutionary steps during the Pleistocene (2.0 to 0.3 Ma) by early members of the genus Homo: H. habilis, H. ergaster, H. erectus; H. heidelbergensis and the earliest H. sapiens. Members of the last three migrated to Eurasia, beginning around 1.8 Ma with the individuals found at Dmanisi in Georgia. The earliest African hominins emerged through the Late Miocene (7.0 to 5.3 Ma): Sahelanthropus tchadensi, Orrorin tugenensis and Ardipthecus kadabba. Through the Pliocene (5.3 to 2.9 Ma) and earliest Pleistocene two very distinct hominin groups appeared: the ‘gracile’ australopithecines (Ardipithecus ramidus; Australopithecus anamensis; Au. afarensis; Au. africanus; Au. sediba) and the ‘robust’ paranthropoids (Paranthropus aethiopicus; P. robustus and P. boisei). The last of the paranthropoids cohabited East Africa with early homo species until around 1.4 Ma. Most of these species have been covered in Earth-logs and an excellent time line of most hominin and early human fossils is hosted by Wikipedia.
All apes, including ourselves, and fossil examples are members of the Family Hominidae (hominids) which refers to the entire world. A Subfamily (Homininae) refers to African apes, with two Tribes. One, the Gorillini, refers to the two living species of gorilla. The other is the Hominini (hominins) that includes chimpanzees, living humans and all fossils believed to be on the evolutionary line to Homo. The Tribe Hominini is defined to have descended from the common ancestor of modern humans and chimps, and evolved only in Africa. As the definition of hominins stands, it excludes other possibilities! The Miocene of Africa before 7.2 Ma ‘goes cold’ as regards the evolution of hominins. There are, however fossils of other African apes in earlier Miocene strata (8 to 18 Ma) that have been assigned to the Family Hominidae, i.e. hominids, of which more later.
Much has been made of using a ‘molecular clock’ to hint at the length of time since the mtDNA of living humans and chimps began to diverge from their last common ancestor. That is a crude measure at it depends entirely on assuming a fixed rate at which genetic mutation in primates take place. Many factors render it highly uncertain, until ancient DNA is recovered from times before about 400 ka, if ever. The approach suggests a range from 7 to 10 Ma, yet the evolutionary history of chimps based on fossils is practically invisible: the earliest fossil of a member of genus Pan is from the Middle Pleistocene (1.2 to 0.8 Ma) of Kenya. Indeed, we have little if any clue about what such a common ancestor looked like or did. So the course of human evolution relies entirely on the fossil sequence of earlier African hominins and comparing their physical appearances. Each species in the African time line displays two distinctive features. All were bipedal and had small canine teeth. Modern chimps habitually use knuckle walking except when having to cross waterways. As with virtually all other primates, fossil or living, male chimps have large, threatening canines. In the absence of ancient DNA from fossils older than 0.4 Ma these two features present a practical if crude way of assessing to when and where the hominin time line leads.
In 2002 a Polish geologist on holiday at the beach at Trachilos on Crete discovered a trackway on a bedding plane in shallow-marine Miocene sediments. It had been left by what seems to have been a bipedal hominin. Subsequent research was able to date the footprints to about 6.05 Ma. Though younger than Sahelanthropus, the discovery potentially challenges the exclusivity of hominins to Africa. Unsurprisingly, publication of this tentative interpretation drew negative responses from some quarters. But the discovery helped resurrect the notion that Africa may have been colonised in the Miocene by hominins that had evolved in Europe. That had been hinted at by the 1872 excavation of Oreopithecus bamboliifrom an Upper Miocene (~7.6 Ma) lignite mine in Tuscany, Italy – a year after publication of Darwin’s The Descent of Man.
Lignites in Tuscany and Sardinia have since yielded many more specimens, so the species is well documented. Oreopithecus could walk on two legs, its hands were capable of a precision grip and it had relatively small canines. Its Wikipedia entry cautiously refers to it as ‘hominid’ – i.e. lumped with all apes to comply with current taxonomic theory (above). In 2019 another fascinating find was made in a clay pit in Bavaria, Germany. Danuvius guggenmosi lived 11.6 Ma ago and fossilised remains of its leg- and arm bones suggested that it could walk on two legs: it too may have been on the hominin line. But no remains of Danuvius’s skull or teeth have been found. There is now an embarrassment of riches as regards Miocene fossil apes from Europe and the Eastern Mediterranean (Sevim-Erol, A. and 8 others 2023. A new ape from Türkiye and the radiation of late Miocene hominines. Nature Communications Biology, v. 6, article 842.; DOI: 10.1038/s42003-023-05210-5). A number of them closely resemble the earliest fossil hominins of Africa, but most predate the hominin record there by several million years.
Phylogenetic links between fossils assigned to Hominidae found in Africa and north of the Mediterranean Sea. (Credit: Sevim-Erol et al. 2023, Fig 5)
Ayla Sevim-Erol of Ankara University, Turkiye and colleagues from Turkiye, Canada and the Netherlands describe a newly identified Miocene genus, Anadoluvius, which they place in the Subfamily Homininae dated to around 8.7 Ma. Fragments of crania and partial male and female mandibles from Anatolia show that its canines were small and comparable with those of younger African hominins, such as Ardipithecus and Australopithecus. But limb bones are yet to be found. Around the size of a large male chimpanzee, Anadoluvius lived in an ecosystem remarkably like the grasslands and dry forests of modern East Africa, with early species of giraffes, wart hogs, rhinos, diverse antelopes, zebras, elephants, porcupines, hyenas and lion-like carnivores. Sevim-Erol et al. have attempted to trace back hominin evolution further than is possible with African fossils. They compare various skeletal features of different fossils and living genera to assess varying degrees of similarity between each genus, applied to 23 genera. These comprised 7 hominids from the African Miocene, 2 early African hominins (Ardipithecus and Orrorin) and 10 Miocene hominids from Europe and the Eastern Mediterranean. They also assessed similarities with 4 living genera, Homo, orang utan (Pongo), gorilla and chimp (Pan).
The resulting phylogeny shows close morphological links within a cluster (green ‘pools’ on diagram) of non-African hominids with the African hominins, gorillas, humans and chimps. There are less-close relations between that cluster and the earlier Miocene hominids of Africa (blue ‘pool’) and the possible phylogeny of orang utans (orange ‘pool’). Sevim-Erol et al. note that African hominins are clearly more similar and perhaps more closely related to the fossils of Europe and the Eastern Mediterranean than they are to Miocene African hominids. This suggests that evolution among the non-African hominids ceased around the end of the Miocene Epoch north of the Mediterranean Sea. But it may have continued in Africa. Somehow, therefore, it became possible late in Miocene times for hominids to migrate from Europe to Africa. Yet the earlier, phylogenetically isolated African hominids seem to have ‘crashed’ at roughly the same time. Such a complex scenario cannot be supported by phylogenetic studies alone: it needs some kind of ecological impetus.
The Mediterranean Basin at the end of the Miocene Epoch when the only water was in the deepest parts of the basin. (Credit: Wikipedia, Creative Commons)
Following a ‘mild’ tectonic collision between the African continent and the Iberian Peninsula during the late Miocene connection between the Atlantic Ocean and the Mediterranean Sea was blocked from 6.0 to 5.3 Ma. Except for its deepest parts, seawater in the Mediterranean evaporated away to leave thick salt deposits. Rivers, such as the Rhône, Danube, Dneiper and Nile, shed sediments into the exposed basin. For 700 ka the basin was a fertile, sub-sea level plain, connecting Europe and North Africa over and E-W distance of 3860 km. There was little to stop the faunas of Eurasia and Africa migrating and intermingling, at a critical period in the evolution of the Family Hominidae. One genus presented with the opportunity was quite possibly the last common ancestor of all the hominins and chimps. The migratory window vanished at the end of the Miocene when what became the Strait of Gibraltar opened at 5.3 to allow Atlantic water. This resulted in the stupendous Zanclean flood with a flow rate about 1,000 times that of the present-day Amazon River. An animation of these events is worth watching
Two decades ago the world of palaeoanthropologists was in turmoil with the publication of an account of a new find in Chad (see: Bonanza time for Bonzo; July 2002). A fossil cranium, dubbed Sahelanthropus tchadensis (nicknamed Toumaï or ‘hope of life’ in the Goran language), appeared like a cross between a chimpanzee and an australopithecine. The turmoil erupted partly because of its age: Upper Miocene, around 7 Ma old. Such an antiquity was difficult to reconcile with the then accepted ~5 Ma estimate for the evolutionary split between humans and chimpanzees, based on applying a ‘molecular clock’ approach to the difference between their mtDNA. The other point of contention was the size of Sahelanthropus’s canine teeth: far too large for australopithecines and humans, but more appropriate for a gorilla or chimp.
Cast of the reconstructed skull of Sahelanthropus tchadensis. (Credit: Didier Descouens, University of Toulouse)
In the absence of pelvic- and foot bones, or signs of the foramen magnum where the spinal cord enters the skull – crucial in distinguishing habitual bipedalism or being an obligate quadruped – encouraged the finders of a 6.1 to 5.7 Ma-old Kenyan hominin Orrorin tugenensis to insist that its skeletal remains – several teeth, fragments of a lower jaw, a thigh bone, an upper arm and of a finger and thumb but no cranial bones – were of ‘the earliest human ancestor’. In Orrorin’s favour were smaller canine teeth than those of later australopithecines. At the time of the dispute, centred mainly on absence of crucial evidence, doyen of hominin fossils Bernard Wood of George Washington University and an advocate of ‘untidy’ evolution, suggested that both early species may well have been evolutionary ‘dead ends’ (see: A considered view; October 2002). And there the ‘muddle’ has rested for 20 years.
In 2002 not only a cranium of Sahelanthropus had been unearthed. Three lower jaw bones and a collection of teeth suggested that as many as 5 individuals had been fossilised. A partial leg bone (femur) and three from forearms (ulna) cannot definitely be ascribed to Sahelanthropus but, in the absence of evidence of any other putative hominin species, they may well be. It has taken two decades for these remains to be analysed to a standard acceptable to peer review (Daver, G. et al. 2022. Postcranial evidence of late Miocene hominin bipedalism in Chad. Nature v. 608, published online; DOI: 10.1038/s41586-022-04901-z). The authors present convoluted anatomical evidence that Toumaï’s femur, which had been gnawed by a porcupine and lacks joints at both ends, suggesting that it was indeed suited to upright walking. Yet the arm bones hint that it may have been equally comfortable in tree canopies. Yet it does look very like an ape rather than a hominin.
Much the same conclusion has been applied to Australopithecus afarensis, indeed its celebrated representative ‘Lucy’ met her end through falling out of a large tree ~3.2 Ma ago (see: Lucy: the australopithecine who fell to Earth?; September 2016). So, dual habitats may have been adopted by hominins long after they emerged. Yet Au afarensis was capable of trudging through mud as witnessed by the famous footprints at Laetoli in Tanzania. Only around 3 Ma has reasonably convincing evidence for upright walking similar to ours been discovered in Au africanus. The full package of signs from pelvis and foot for habitual bipedalism dates to 2 Ma ago in Au sediba. Even this latest known australopithecine seems to have had a gait oddly different from that of members of the genus Homo.
So, in many respects the benefits of full freeing of the hands to develop manipulation of objects, as first suggested by Freidrich Engels, may have had to await the appearance of early humans. Earlier hominins almost certainly did make tools of a kind, but the revolutionary breakthrough associated with humanity was more than 5 million years in the making.
To see traces of where our forebears walked, such as the famous Australopithecusafarensistrackway at Laetoli in Tanzania, the footprints of Neanderthal children in 350 ka old Italian volcanic ash (The first volcanologists? Earth Pages March 2003) or even those of Mesolithic families in estuarine mud is about as heart stopping as it gets for a geologist. But imagine the astonishment of members of a multinational team working on Miocene shore-line sediments on Crete when they came upon a bedding surface covered with what are almost certainly the footprints of another bipedal animal from 5.7 Ma ago (Gierliński, G.D. et al. 2017. Possible hominin footprints from the late Miocene (c. 5.7 Ma) of Crete? Proceedings of the Geologists’ Association, online; https://doi.org/10.1016/j.pgeola.2017.07.006). Trackways preserve a few moments in time, however old they are and the chances of their being preserved are very small, yet they can supply information that is lost from even the best preserved fossil, such as gait, weight, speed and so forth.
Track bearing surface; (b) two footprints in 5.7 Ma old Miocene sediments at Trachilos, Crete (credit: Gierliński, G.D. et al. 2017; Figures 2 and 8)
The tracks clearly indicate that whatever left them was bipedal and lacked claws, and closely resemble those attributed to A. afarensis at Laetoli in a 3.7 Ma old volcanic ash. What they do not resemble closely are those of non-hominin modern primates, such as chimpanzees. They are diminutive compared with adult modern human prints, being about 12.5 cm long (equivalent to a UK child’ shoe size 4 – US size 4.5, EU 20) and about a third to half the size of those at Laetoli. Were they around the age of those at Laetoli or younger there seems little doubt that they would be widely interpreted as being of hominin origin. But being from an island in the Mediterranean as well as far from sites in Africa that have yielded Miocene hominins (Ardipithecus kadabba from Ethiopia, Orrorin from Kenya and Sahelanthropusfrom Chad), such an interpretation is bound to create controversy. Somewhat less controversial might be to regard them as having been created by a late-Miocene primate that convergently evolved a hominin-like upright gait and foot. Being preserved in what seem to be coastal marine sediments, there is probably little chance of body fossils being preserved in the exposed horizon. Since foot bones are so fragile, even if a primate fossil is discovered in the late Miocene of Crete the chances of resolving the issue are pretty remote. Yet fossil primate specialists will undoubtedly beat a well-trodden path to the Trachilos site near Kissamos on Crete
We know that Lucy, the famous Australopithecus afarensis, could climb trees because her many bone fractures show that she fell out of a tree to her death. But that does not mean her species was an habitual tree-climber: plenty of modern humans fall to their deaths from trees, cliffs and the like. But the issue seems to have been resolved by using X-ray tomography of Lucy’s limb bones (Ruff, C.B. et al. 2016. Limb bone structural proportions and locomotor behaviour in A.L. 288-1 (“Lucy”). PLOS ONE v. 11, e0166095. doi:10.1371/journal.pone.0166095) during the skeleton’s triumphal series of exhibits in the US.
The authors, including two of those who showed that Lucy died after a fall using similar data, compared the digital 3-D models of her surviving arm- and leg bones with those of other hominins and living primates, estimating their relative strengths at different positions. Lucy was probably stronger in the arm than in the leg, but not to the same degree as chimpanzees. This is a feature that would significantlyassist climbing , but her bipedal locomotion on the ground would have been only slightly different from that of later Homo species. If anything, her strength relative to size would have been greater than ours, perhaps reflecting less reliance on tools for getting food and defending herself. But almost certainly Australopithecus afarensis habitually spend more time in trees, perhaps foraging and as a defence against predation, especially at night.
The new data for Lucy allows palaeoanthropologists to better judge the capabilities of other hominins. Interestingly Homo habilis, the earliest of our genus, may have had similar habits. But later species, beginning with H. erectus/ergaster, were as Earth-bound as we are. This suggests a shift in hominin ecology from an early and probably long history of semi-arboreal behavior until humans became masters of their terrain about 1.9 Ma ago, probably through their invention of better tools and the controlled use of fire.




















{kind=link}