Hominins have been making and using stone tools since at least 3.4 Ma, as shown by cut marks on bones and stone artefacts themselves. I use the sack term ‘hominin’ because the likely makers and users of the oldest tools are either australopithecines or paranthropoids, there being no fossils designates to the genus Homo of late-Pliocene age. So it might seem un-newsworthy to report that the oldest tools deliberately made from bone are now known to occur in 1.5 Ma old sediments from the famous sedimentary sequence at Olduvai Gorge in Tanzania (de la Torre, I and 8 others2025. Systematic bone tool production at 1.5 million years ago. Nature, v. 639; DOI: 10.1038/s41586-025-08652-5). To be clear, there is abundant evidence that hominins had used bones, especially sturdy long bones, for digging perhaps, much earlier in hominin history. Again, paranthropoids have been implicated in their use. The bones found at Olduvai actually show signs of manufacture into useful objects prior to their use: they show clear signs of knapping to produce points and blades. The bones are among the sturdiest known from the Pleistocene, being from elephants and hippos. Before de la Torre and colleagues found what is essentially a bone-tool factory, it was thought that systematic use of bones in such a sophisticated manner only arose between 400 to 250 ka ago among early Homo in Europe. Sadly, fossils of whoever made the tools were not found at the site. Once again, paranthropoids as well as early Homo are known to have cohabited the area at that time.
‘Front, back and side’ views of a 1.5 Ma old tool made from an elephant humerus – its upper foreleg. The scale bar represents 5 cm. (Credit: de la Torre et al.; Fig 3a)
Bifacial Acheulean stone artefacts first appear in the rock record about 300 ka before these bone tools were made. So one idea that the authors put forward is that the same kind of stone knapping technique was transferred to the more abundant massive bones of the East African Pleistocene megafauna (in the absence or rarity of suitable blocks of stone?). But it remains unclear whether or not such tools were simply selected from very large bones smashed to get at their nutritious marrow. The first possibility implies a cultural shift, whereas the latter points simply to expedience. The authors are at pains to point out that the curious million-year gap in the record of bone tools may be ascribed either to the disappearance of bone technology or simply to archaeologists who worked elsewhere having not regarding bone fragments as the products of skills. That applies equally to earlier times, when bones were indeed used, though with not so much in the way of a ‘mental template’. As de la Torre et al. conclude ‘Future research needs to investigate whether similar bone tools were already produced in earlier times, persisted during the Acheulean and eventually evolved into Middle Pleistocene bone bifaces similar in shape, size and technology to their stone counterparts’.
Charles Darwin famously suggested that humans evolved from apes, and since great apes (chimpanzees, bonobos and gorillas) live in Africa he reckoned it was probably there that the human ‘line’ began. Indeed, the mitochondrial DNA of chimpanzees (Pan troglodytes) is the closest to that of living humans. Palaeoanthropology in Africa has established evolutionary steps during the Pleistocene (2.0 to 0.3 Ma) by early members of the genus Homo: H. habilis, H. ergaster, H. erectus; H. heidelbergensis and the earliest H. sapiens. Members of the last three migrated to Eurasia, beginning around 1.8 Ma with the individuals found at Dmanisi in Georgia. The earliest African hominins emerged through the Late Miocene (7.0 to 5.3 Ma): Sahelanthropus tchadensi, Orrorin tugenensis and Ardipthecus kadabba. Through the Pliocene (5.3 to 2.9 Ma) and earliest Pleistocene two very distinct hominin groups appeared: the ‘gracile’ australopithecines (Ardipithecus ramidus; Australopithecus anamensis; Au. afarensis; Au. africanus; Au. sediba) and the ‘robust’ paranthropoids (Paranthropus aethiopicus; P. robustus and P. boisei). The last of the paranthropoids cohabited East Africa with early homo species until around 1.4 Ma. Most of these species have been covered in Earth-logs and an excellent time line of most hominin and early human fossils is hosted by Wikipedia.
All apes, including ourselves, and fossil examples are members of the Family Hominidae (hominids) which refers to the entire world. A Subfamily (Homininae) refers to African apes, with two Tribes. One, the Gorillini, refers to the two living species of gorilla. The other is the Hominini (hominins) that includes chimpanzees, living humans and all fossils believed to be on the evolutionary line to Homo. The Tribe Hominini is defined to have descended from the common ancestor of modern humans and chimps, and evolved only in Africa. As the definition of hominins stands, it excludes other possibilities! The Miocene of Africa before 7.2 Ma ‘goes cold’ as regards the evolution of hominins. There are, however fossils of other African apes in earlier Miocene strata (8 to 18 Ma) that have been assigned to the Family Hominidae, i.e. hominids, of which more later.
Much has been made of using a ‘molecular clock’ to hint at the length of time since the mtDNA of living humans and chimps began to diverge from their last common ancestor. That is a crude measure at it depends entirely on assuming a fixed rate at which genetic mutation in primates take place. Many factors render it highly uncertain, until ancient DNA is recovered from times before about 400 ka, if ever. The approach suggests a range from 7 to 10 Ma, yet the evolutionary history of chimps based on fossils is practically invisible: the earliest fossil of a member of genus Pan is from the Middle Pleistocene (1.2 to 0.8 Ma) of Kenya. Indeed, we have little if any clue about what such a common ancestor looked like or did. So the course of human evolution relies entirely on the fossil sequence of earlier African hominins and comparing their physical appearances. Each species in the African time line displays two distinctive features. All were bipedal and had small canine teeth. Modern chimps habitually use knuckle walking except when having to cross waterways. As with virtually all other primates, fossil or living, male chimps have large, threatening canines. In the absence of ancient DNA from fossils older than 0.4 Ma these two features present a practical if crude way of assessing to when and where the hominin time line leads.
In 2002 a Polish geologist on holiday at the beach at Trachilos on Crete discovered a trackway on a bedding plane in shallow-marine Miocene sediments. It had been left by what seems to have been a bipedal hominin. Subsequent research was able to date the footprints to about 6.05 Ma. Though younger than Sahelanthropus, the discovery potentially challenges the exclusivity of hominins to Africa. Unsurprisingly, publication of this tentative interpretation drew negative responses from some quarters. But the discovery helped resurrect the notion that Africa may have been colonised in the Miocene by hominins that had evolved in Europe. That had been hinted at by the 1872 excavation of Oreopithecus bamboliifrom an Upper Miocene (~7.6 Ma) lignite mine in Tuscany, Italy – a year after publication of Darwin’s The Descent of Man.
Lignites in Tuscany and Sardinia have since yielded many more specimens, so the species is well documented. Oreopithecus could walk on two legs, its hands were capable of a precision grip and it had relatively small canines. Its Wikipedia entry cautiously refers to it as ‘hominid’ – i.e. lumped with all apes to comply with current taxonomic theory (above). In 2019 another fascinating find was made in a clay pit in Bavaria, Germany. Danuvius guggenmosi lived 11.6 Ma ago and fossilised remains of its leg- and arm bones suggested that it could walk on two legs: it too may have been on the hominin line. But no remains of Danuvius’s skull or teeth have been found. There is now an embarrassment of riches as regards Miocene fossil apes from Europe and the Eastern Mediterranean (Sevim-Erol, A. and 8 others 2023. A new ape from Türkiye and the radiation of late Miocene hominines. Nature Communications Biology, v. 6, article 842.; DOI: 10.1038/s42003-023-05210-5). A number of them closely resemble the earliest fossil hominins of Africa, but most predate the hominin record there by several million years.
Phylogenetic links between fossils assigned to Hominidae found in Africa and north of the Mediterranean Sea. (Credit: Sevim-Erol et al. 2023, Fig 5)
Ayla Sevim-Erol of Ankara University, Turkiye and colleagues from Turkiye, Canada and the Netherlands describe a newly identified Miocene genus, Anadoluvius, which they place in the Subfamily Homininae dated to around 8.7 Ma. Fragments of crania and partial male and female mandibles from Anatolia show that its canines were small and comparable with those of younger African hominins, such as Ardipithecus and Australopithecus. But limb bones are yet to be found. Around the size of a large male chimpanzee, Anadoluvius lived in an ecosystem remarkably like the grasslands and dry forests of modern East Africa, with early species of giraffes, wart hogs, rhinos, diverse antelopes, zebras, elephants, porcupines, hyenas and lion-like carnivores. Sevim-Erol et al. have attempted to trace back hominin evolution further than is possible with African fossils. They compare various skeletal features of different fossils and living genera to assess varying degrees of similarity between each genus, applied to 23 genera. These comprised 7 hominids from the African Miocene, 2 early African hominins (Ardipithecus and Orrorin) and 10 Miocene hominids from Europe and the Eastern Mediterranean. They also assessed similarities with 4 living genera, Homo, orang utan (Pongo), gorilla and chimp (Pan).
The resulting phylogeny shows close morphological links within a cluster (green ‘pools’ on diagram) of non-African hominids with the African hominins, gorillas, humans and chimps. There are less-close relations between that cluster and the earlier Miocene hominids of Africa (blue ‘pool’) and the possible phylogeny of orang utans (orange ‘pool’). Sevim-Erol et al. note that African hominins are clearly more similar and perhaps more closely related to the fossils of Europe and the Eastern Mediterranean than they are to Miocene African hominids. This suggests that evolution among the non-African hominids ceased around the end of the Miocene Epoch north of the Mediterranean Sea. But it may have continued in Africa. Somehow, therefore, it became possible late in Miocene times for hominids to migrate from Europe to Africa. Yet the earlier, phylogenetically isolated African hominids seem to have ‘crashed’ at roughly the same time. Such a complex scenario cannot be supported by phylogenetic studies alone: it needs some kind of ecological impetus.
The Mediterranean Basin at the end of the Miocene Epoch when the only water was in the deepest parts of the basin. (Credit: Wikipedia, Creative Commons)
Following a ‘mild’ tectonic collision between the African continent and the Iberian Peninsula during the late Miocene connection between the Atlantic Ocean and the Mediterranean Sea was blocked from 6.0 to 5.3 Ma. Except for its deepest parts, seawater in the Mediterranean evaporated away to leave thick salt deposits. Rivers, such as the Rhône, Danube, Dneiper and Nile, shed sediments into the exposed basin. For 700 ka the basin was a fertile, sub-sea level plain, connecting Europe and North Africa over and E-W distance of 3860 km. There was little to stop the faunas of Eurasia and Africa migrating and intermingling, at a critical period in the evolution of the Family Hominidae. One genus presented with the opportunity was quite possibly the last common ancestor of all the hominins and chimps. The migratory window vanished at the end of the Miocene when what became the Strait of Gibraltar opened at 5.3 to allow Atlantic water. This resulted in the stupendous Zanclean flood with a flow rate about 1,000 times that of the present-day Amazon River. An animation of these events is worth watching
The largest volcanic eruption during the 2.5 million year evolution of the genius Homo, about 74 thousand years (ka) ago, formed a huge caldera in Sumatra, now filled by Lake Toba. A series of explosions lasting just 9 to 14 days was forceful enough to blast between 2,800 to 6,000 km3 of rocky debris from the crust. An estimated 800 km3 was in the form of fine volcanic ash that blanketed South Asia to a depth of 15 cm. Thin ash layers containing shards of glass from Toba occur in marine sediments beneath the Indian Ocean, the Arabian and South China Seas. Some occur as far off as sediments on the floor of Lake Malawi in southern Africa. A ‘spike’ of sulfates is present at around 74 ka in a Greenland ice core too. Stratospheric fine dust and sulfate aerosols from Toba probably caused global cooling of up to 3.5 °C over a modelled 5 years following the eruption. To make matters worse, this severe ‘volcanic winter’ occurred during a climatic transition from warm to cold caused by changes in ocean circulation and falling atmospheric CO2 concentration, known as a Dansgaard-Oeschger event.
There had been short-lived migrations of modern humans out of Africa into the Levant since about 185 ka. However, studies of the mitochondrial DNA (mtDNA) of living humans in Eurasia and Australasia suggest that permanent migration began about 60 ka ago. Another outcome of the mtDNA analysis is that the genetic diversity of living humans is surprisingly low. This suggests that human genetic diversity may have been sharply reduced globally roughly around the time of the Toba eruption. This implies a population bottleneck with the number of humans alive at the time to the order of a few tens of thousands (see also: Toba ash and calibrating the Pleistocene record; December 2012). Could such a major genetic ‘pruning’ have happened in Africa? Over six field seasons, a large team of geoscientists and archaeologists drawn from the USA, Ethiopia, China, France and South Africa have excavated a rich Palaeolithic site in the valley of the Shinfa River, a tributary of the Blue Nile in western Ethiopia. Microscopic studies of the sediments enclosing the site yielded glass shards whose chemistry closely matches those in Toba ash, thereby providing an extremely precise date for the human occupation of the site: during the Toba eruption itself (Kappelman, Y. and 63 others 2024. Adaptive foraging behaviours in the Horn of Africa during Toba supereruption. Nature, v. 627; DOI: 10.1038/s41586-024-07208-3).
Selection of possible arrowheads from the Shinfa River site (Credit: Kappelman et al.; Blue Nile Survey Project)
The artifacts and bones of what these modern humans ate suggest a remarkable scenario for how they lived. Stone tools are finely worked from local basalt lava, quartz and flint-like chalcedony found in cavities in lava flows. Many of them are small, sharp triangular points, some of which show features consistent with their use as projectile tips that fractured on impact; they may be arrowheads, indeed the earliest known. Bones found at the site are key pointers to their diet. They are from a wide variety of animal, roughly similar to those living in the area at present: from monkeys to giraffe, guinea fowl to ostrich, and even frogs. There are remains of many fish and freshwater molluscs. Although there are no traces of plant foods, clearly those people who loved through the distant effects of Toba were well fed. Although a period of global cooling may have increased aridity at tropical latitudes in Africa, the campers were able to devise efficient strategies to obtain victuals. During wet seasons they lived off terrestrial prey animals, and during the driest times ate fish from pools in the river valley. These are hardly conditions likely to devastate their numbers, and the people seem to have been technologically flexible. Similar observations were made at the Pinnacle Point site in far-off South Africa in 2018, where Toba ash is also present. Both sites refute any retardation of human cultural progress 74 ka ago. Rather the opposite: people may have been spurred to innovation, and the new strategies may have allowed them to migrate more efficiently, perhaps along seasonal drainages. In this case that would have led them or their descendants to the Nile and a direct route to Eurasia; along ‘blue highway’ corridors as Kappelman et al. suggest.
Yet the population bottleneck implied by mtDNA analyses is only vaguely dated: it may have been well before or well after Toba. Moreover, there is a 10 ka gap between Toba and the earliest accurately dated migrants who left Africa – the first Australians at about 65 ka. However, note that there is inconclusive evidence that modern humans may have occupied Sumatra by the time of the eruption. Much closer to the site of the eruption in southeast India, stone artifacts have been found below and above the 74 ka datum marked by the thick Toba Ash. Whether these were discarded by anatomically modern humans or earlier migrants such as Homo erectus remains unresolved. Either way, at that site there is no evidence for any mass die-off, even though conditions must have been pretty dreadful while the ash fell. But that probably only lasted for little more than a month. If the migrants did suffer very high losses to decrease the genetic diversity of the survivors, it seems just as likely to have been due to attrition on an extremely lengthy trek, with little likelihood of tangible evidence surviving. Alternatively, the out-of-Africa migrants may have been small in number and not fully representative of the genetic richness of the Africans who stayed put: a few tens of thousand migrants may not have been very diverse from the outset.
Reconstruction of a Paranthropus head (Credit: Jerry Humphrey, Pinterest)
Paranthropoids had large, broad teeth and pronounced cheekbones plus a bone crest on the top of their skulls that were the attachments for powerful jaw muscles, much as in modern gorillas. Unlike gorillas they were definitely bipedal and were more similar to australopithecines. They have been called ‘robust’ australopithecines but they were not significantly taller or heavier. The first to be unearthed at Olduvai, Tanzania in 1959 (Paranthropus boisei) was dubbed ‘Nutcracker Man’ by its finder, and many have implied that paranthropoids’ teeth and powerful jaws were signs of a vegetarian diet that needed a lot of chewing. Yet their teeth do not show the microscopic pitting associated with living primates that eat hard plant parts and nuts, or the heavy wear that results from eating grasses. They probably ate soft plants, such as semi-aquatic succulents or tubers, but meat-eating that causes little dental wear cannot be ruled out. Some specimens are associated with long bones of other animals whose ends are worn, suggesting that they may have used them as tools for digging. Plant remains found at paranthropoid sites suggests that they inhabited woodland, together with coexisting australopithecines. They were around in the form of three successive species from 2.9 to 1.2 Ma, outlasting australopithecines. The later paranthropoids coexisted with Homo habilis and H. erectus: they were clearly just as successfully adapted to their surroundings as were early humans.
In early 2023 evidence was published that associated Oldowan stone tools with remains of Paranthropus, together with deliberately defleshed and cut bones (see also): though association is not proof of a direct link. Interestingly, the hand of a P. robustus found in the Swartkrans cave system in South Africa is consistent with a human-like precision grip, i.e. it had an opposable thumb. Swarkrans also yielded the earliest evidence for the deliberate use of fire about 1.5 Ma ago, associated with remains of both P. robustus and H. erectus. All this suggests that a case could be made for paranthropoids’ being human ancestors – supporting evidence has just been published (Braga, J. et al. 2023. Hominin fossils from Kromdraai and Drimolen inform Paranthropus robustus craniofacial ontogeny. Science Advances, v. 9, article eade7165; DOI: 10.1126/sciadv.ade7165).
Fossil-bearing breccias beneath the floor of the Kromdraai cave in the Cradle of HumankindWorld Heritage Site 45 km NW of Johannesburg, South Africa yielded the first near-complete P. robustus skull in 1938, another being found in cave breccias at the nearby Drimolen quarry. These deposits also contained remains of four infants assigned to the species, whose teeth and cranial parts were at different stages of juvenile development (ontogeny). José Braga of the University of Toulouse, France and co-workers from South Africa and the USA compared this growth sequence with those teased out from immature specimens of Australopithecus africanus and early Homo.Their tentative conclusion is that Paranthropus robustus is more closely related to early humans than to australopithecines of the same stratigraphic age.
Skull of a probable adult female P. robustus (left) with that of H. habilis (centre) and A. africanus (right). Credits: all from Wikipedia pages
So, it now seems possible that paranthropoids are not ‘robust’ australopithecines in an acceptable, taxonomic sense. Their closer resemblance in early development to early humans, together with their association with early stone tools used for defleshing prey animals, together with evidence for possible their use of fire, further strengthens their candidacy for an ancestral link to humans. The best preserved skulls of Homo habilis and a female P. robustus (males of that species show the distinctive saggital crest) do show close similarities, that of a roughly contemporary A. africanus having distinctly wider cheeks than both. All three species were in life probably of much the same weight and stature (30 to 40 kg and 110 to 130 cm) but H. habilis had a significantly larger brain volume (500 to 900 cm3) than the other two (each ~450 cm3). However, this isn’t proof that the genus Homo evolved from a paranthropoid ancestor. That would require genetic evidence, unlikely to be extracted from specimens because DNA seems to degrade more quickly under the conditions of the tropics than at high latitudes. Debate on ultimate human origins will probably be endless. Perhaps it would make more sense simply to accept that early humans weren’t the only ‘smart kids on the palaeoanthropological block’, one of which left no issue after 1.2 Ma ago.
Steadily, the record of stone tools has progressed further back in time as archaeological surveys have expanded, especially in East Africa (Stone tools go even further back, May 2015). The earliest known tools – now termed Lomekwian – are 3.3 million years old, from deposits in north-western Kenya, as are cut-marked bone fragments from Ethiopia’s Afar region. There is no direct link to their makers, but at least six species of Australopithecus occupied Africa during the Middle Pliocene. Similarly, there are various options for who made Oldowan tools in the period between 2.6 and 2.0 Ma, the only known direct association being with Homo habilis in 2.0 Ma old sediments from Tanzania’s Olduvai Gorge; the type locality for the Oldowan.
The shapes of stone tools and the manufacturing techniques required to make them and other artefacts, are among the best, if not the only, means of assessing the cognitive abilities of their makers. A new, detailed study of the shapes of 327 Oldowan tools from a 2.6 Ma old site in Afar, Ethiopia has revealed a major shift in hominin working methods (Braun, D.R. and 17 others 2019. Earliest known Oldowan artifacts at >2.58 Ma from Ledi-Geraru, Ethiopia, highlight early technological diversity. Proceedings of the National Academy, v. 116, p. 11712-11717; DOI: 10.1073/pnas.1820177116). The sharp-edged tools were made by more complex methods than the Lomekwian. Analysis suggests that they were probably made by striking two lumps of rock together, i.e. by a deliberate two-handed technique. On the other hand, Lomekwian tools derived simply by repeatedly bashing one rock against a hard surface, not much different from the way some living primates make rudimentary tools. But the morphology of the Ledi-Geraru tools also falls into several distinct types, each suggesting systematic removal of only 2 or 3 flakes to make a sharp edge. The variations in technique suggest that several different groups with different traditions used the once lake-side site.
Various 2.6 Ma old Oldowan stone tools from Ledi-Geraru, Ethiopia (credit: Braun et al., 2019)
Ledi-Geraru lies about 5 km from another site dated about 200 ka earlier than the tools, which yielded a hominin jawbone, likely to be from the earliest known member of the genus Homo. A key feature that suggested a human affinity is the nature of the teeth that differ markedly from those of contemporary and earlier australopithecines. It appears that the tools are of early human manufacture. The ecosystem suggested by bones of other animals, such as antelope and giraffe was probably open grassland – a more difficult environment for hominin subsistence. The time of the Lomekwian tools was one of significantly denser vegetation, with more opportunities for gathering plant foods. Perhaps this environmental shift was instrumental in driving hominins to increased scavenging of meat, the selection pressure acting on culture to demand tools sharp enough to remove meat from the prey of other animals quickly, and on physiology and cognitive power to achieve that.
Bill Deller, a friend whose Sunday is partly spent reading the Observer and Sunday Times from cover to cover, alerted me to a lengthy article by Britain’s doyen of paleoanthropologists Chris Stringer of the Natural History Museum. (Stringer, C. 2015. First human? The jawbone that makes us question where we’re from. Observer, 8 March 2015, p. 36). His piece sprang from two Reports published online in Science that describe about 1/3 of a hominin lower jaw unearthed – where else? – in the Afar Depression of Ethiopia. The discovery site of Ledi-Geraru is a mere 30 km from the most hominin-productive ground in Africa: Hadar and Dikika for Australopithecus afarensis (‘Lucy’ at 3.2 Ma and ‘Selam’ at 3.3 Ma, respectively); Gona for the earliest-known stone tools (2.6 Ma); and the previously earliest member of the genus Homo, also close to Hadar.
On some small objects mighty tales are hung, and the Ledi-Geraru jawbone and 6 teeth is one of them. It has features intermediate between Australopithecus and Homo, but more important is its age: Pliocene, around 2.8 to 2.75 Ma (Villmoare, B. And 8 others. Early Homo at 2.8 Ma from Ledi Geraru, Afar, Ethiopia. Science Express doi: 10.1126/science.aaa1343). The sediments from which Ethiopian geologist Chalachew Seyoum, studying at Arizona State University, extracted the jawbone formed in a river floodplain. Other fossils suggest open grassland rich with game, similar to that of the Serengeti in Tanzania, with tree-lined river courses. These were laid down at a time of climatic transition from humid to more arid conditions, that several authors have suggested to have provided the environmental stresses that drove evolutionary change, including that of hominins (DiMaggio, E.N. and 10 others 2015. Late Pliocene fossiliferous sedimentary record and the environmental context of early Homo from Afar, Ethiopia. Science Express doi: 10.1126/science.aaa1415).
Designating the jawbone as evidence for the earliest known member of our genus rests almost entirely on the teeth, and so is at best tentative awaiting further fossil material. The greatest complicating factor is that the earliest supposed fossils of Homo (i.e. H. habilis, H rudolfensis and others yet to be assigned a species identity) are a morphologically more mixed bunch than those younger than 2 Ma, such as H. ergaster and H. erectus. Indeed, every one of them has some significant peculiarity. That diversity even extends to the earliest humans to have left Africa, found in 1.8 Ma old sediments at Dmanisi in Georgia (Homo georgicus), where each of the 5 well-preserved skulls is unique. The Dmanisi hominins have been likened to the type specimen of H. habilis, but such is the diversity of both that is probably a shot in the dark.
Replica of OH 7, the deformed type specimen of Homo habilis. (credit: Wikipedia)
Coinciding with the new Ethiopian hominin papers a study was published in Nature the same week that describes how the type specimen of H. habilis (found, in close association with crude stone tools and cut bones, by Mary and Lewis Leakey at Olduvai Gorge, Tanzania in 1960) has been digitally restored from its somewhat deformed state when found (Spoor, F. et al. 2015. Reconstructed Homo habilis type OH 7 suggests deep-rooted species diversity in early Homo. Nature, v. 519, p. 83-86, doi:10.1038/nature14224). The restored lower jaw and teeth, and part of its cranium, deepened the mysterious diversity of the group of fossils for which it is the type specimen, but boosts its standing as regards probable brain size from one within the range of australopithecines to significantly larger –~750 ml compared with <600 ml – about half that of modern humans. The habilis diversity is largely to do with jaws and teeth: it is the estimated brain size as well as the type specimen’s association with tools and their use that elevates them all to human status. Yet, the reconstruction is said by some to raise the issue of a mosaic of early human species. The alternative is an unusual degree of shape diversity (polymorphism) among a single emerging species, which is not much favoured these days. An issue to consider is: what constitutes a species? For living organisms morphological similarity has to be set against the ability for fertile interbreeding. Small, geographically isolated populations of a single species often diverge markedly in terms of what they look like yet continue to be interfertile, the opposite being convergence in form by organisms that are completely unrelated.
Palaeontologists tend to go largely with division on grounds of form, so that when a specimen falls outside some agreed morphological statistics, it crosses a species boundary. Set against that the incontrovertible evidence that at least 3 recent human species interbred successfully to leave the mark in all non-African living humans. What if the first humans emerging from, probably, a well-defined population of australopithecines continued to interbreed with them, right up to the point when they became extinct about 2 Ma ago?
On a more concrete note, the Ledi Geraru hominin is a good candidate for the maker of the first stone tools found ‘just down the road’ at Gona!
A press release from the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, announces the completion of the most precise genome from a third Neanderthal individual. For the first time it is possible to distinguish copies of the genes inherited by the individual from both parents. The data release coincided with a review of genetic evidence for interbreeding between early Homo sapiens and other species.
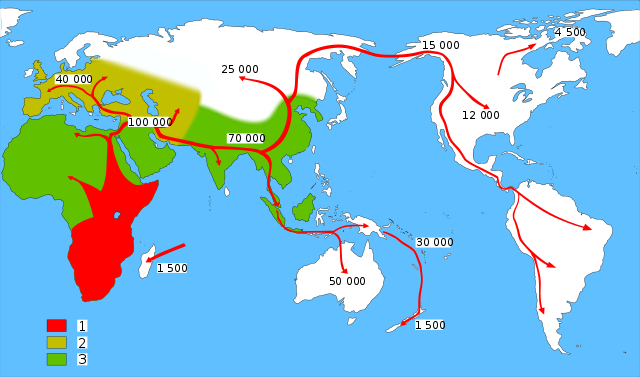
Modern human migration out of and within Africa relative to the domains of coeval archaic humans 1 = modern humans 2 = Neanderthals 3 = other archaic humans (credit: Wikipedia)