
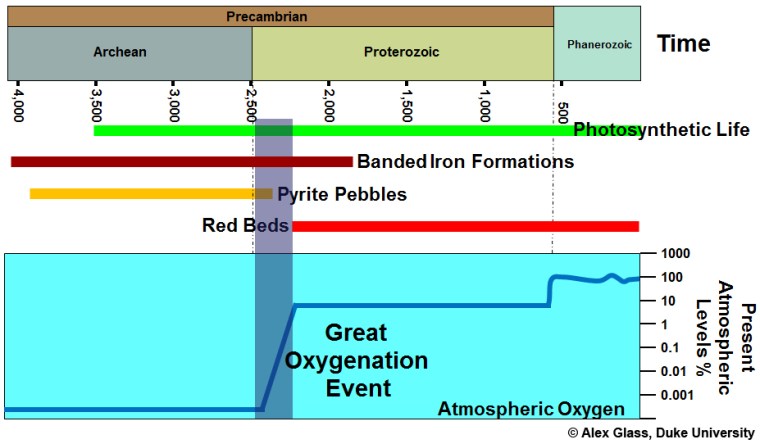
Earth’s present atmosphere contains oxygen because of one form of photosynthesis that processes water and carbon dioxide to make plant carbohydrates, leaving oxygen at a waste product. The photochemical trick that underpins oxygenic photosynthesis seems only to have evolved once. It was incorporated in a simple, single-celled organism or prokaryote, which lacks a cell nucleus but contains the necessary catalyst chlorophyll. Such an organism gave rise to cyanobacteria or blue-green bacteria, which still make a major contribution to replenishing atmospheric oxygen. Chloroplasts that perform the same function in plant cells are so like cyanobacteria that they were almost certainly co-opted during the evolution of a section of nucleus-bearing eukaryotes that became the ancestors of plants. A range of evidence suggests that oxygenic photosynthesis appeared during the Archaean Eon, the most tangible being the presence of stromatolites, which cyanobacteria mats or biofilms form today. These knobbly structures in carbonate sediments extend as far back as 3.5 billion years ago (see: Signs of life in some of the oldest rocks; September 2016). Yet it took a billion years before the first inklings of biogenic oxygen production culminated in the Great Oxygenation Event or GOE (see: Massive event in the Precambrian carbon cycle; January, 2012) at around 2400 Ma. Then, for the first time, oxidised iron in ancient soils turned them red. If oxygen was being produced, albeit in small amounts, in shallow, sunlit Archaean seas, why didn’t it build up in the atmosphere of those times? Geochemical analyses of Archaean sediments do point to trace amounts, with a few ‘whiffs’ of more substantial amounts. But they fall well below those of Meso- and Neoproterozoic and Phanerozoic times. One hypothesis is that Archaean oceans contained dissolved, ferrous iron (Fe2+) – a powerful reducing agent – with which available oxygen reacted to form insoluble ferric iron (Fe3+) oxides and hydroxides that formed banded iron formations (BIFS). The Fe2+ in this hypothesis is attributed to hydrothermal activity in basaltic oceanic crust. There is, however, another possibility for suppression of atmospheric oxygen accumulation in the Archaean and early-Palaeoproterozoic.
Simone Marchi of the Southwest Research Institute of Boulder, CO, USA and colleagues from the US, Austria and Germany suggest that planetary bombardment offers a plausible explanation (Marchi, S. et al 2021. Delayed and variable late Archaean atmospheric oxidation due to high collision rates on Earth. Nature Geoscience, v. 14 advance publication; DOI: 10.1038/s41561-021-00835-9). Over the last 20 years evidence of extraterrestrial impacts has emerged, in the form of thin spherule-bearing layers in Archaean sedimentary strata, probably formed by impacts of objects around 10 km across. So far 35 such layers have been identified from several locations in South Africa and Western Australia. They span the last billion years of the Archaean and the earliest Palaeoproterozoic, although they are not evenly spaced in time. The spherules represent droplets of mainly crustal but some meteoritic rocks that were vaporised by impacts and then condensed as liquid. Meteorites in particular contain reduced elements and compounds, including iron, whose oxidation by would remove free oxygen.
The evidence from spherule beds is supplemented by the team’s new calculations of the likely flux of impactors during the Archaean. These stem from re-evaluation of the lunar cratering record that is used to estimate the number and size of impacts on Earth up to 2.5 Ga ago. This flux amounts to the ‘leftovers’ of the catastrophic period around 4.1 Ga when the giant planets Jupiter and Saturn ran amok before they settled into their present orbits. Their perturbation of gravitational fields in the solar system injected a long-lived supply of potential impactors into the inner solar system, which is recorded by craters on the post-4.1 Ga lunar maria. The calculations suggest that the known spherule layers underestimate the true number of such collisions on Earth. Modelling by Marchi et al., based on the meteorite flux and the oxidation of vaporised materials produced by impacts, plausibly accounts for the delay in atmospheric oxygen build-up.
It is worth bearing in mind, however, that large impacts and their geochemical aftermath are, in a geological sense, instantaneous events widely spaced in time. They may have chemically ‘sucked’ oxygen out of the Archaean and early-Palaeoproterozoic atmosphere. Yet photosynthesising bacteria would have been generating oxygen continuously between such sudden events. The same goes for the supply of reduced ferrous iron and its circulation in the oceans of those times, capable of scavenging available oxygen through simple chemical reactions. In fact we can still observe that in action around ocean-floor hydrothermal vents where a host of reduced elements and compounds are oxidised by dissolved oxygen. The difference is that oxygen is now produced more efficiently on land and in the upper oceans and a less vigorous mantle is adding less iron-rich basalt magma to the crust: the balance has changed. Another issue is that the Great Oxygenation Event terminated the oxygen-starved conditions of the Archaean and Palaeoproterozoic in about 200 million years, despite the vast production of BIFs before and after it happened. The Wikipedia entry for the GOE provides a number of hypotheses for how that termination came about. Interestingly, one idea looks to a shortage of dissolved nickel that is vital for methane generating bacteria: a nickel ‘famine’. A geochemical setback for methanogens would have been a boost for oxygenic photosynthesisers and especially their waste product oxygen: methane quickly reacts with oxygen in the atmosphere to produce CO2 and water. Anomalously high nickel is a ‘signature element’ for meteorite bombardment, though it can be released by hydrothermal alteration of basalt. Had meteoritic nickel been fertilising methane-generating bacteria in the oceans prior to the GOE?
See also: A new Earth bombardment model. Science Daily, 21 October 2021.



