
Humans are more or less symmetrical, our left and right sides closely resembling each other. That is not so comprehensive for our innards, except for testes and ovaries, kidneys, lungs, arteries and veins, lymph and nervous systems. We have front- and rear ends, top and bottom, input and output orifices. All that we share with almost all other animals from mammals to worms, particularly at the earliest, embryonic stage of development. We are bilaterians, whereas sponges, ctenophores, placozoans and cnidarians are not – having either no symmetry at all, or just a bottom and a top – and are in a minority. Fossil collections from Cambrian times also reveal bilaterians in the majority, at least insofar as preservation allows us to tell. Before 541 Ma ago, in the Precambrian, there are few signs of such symmetry and faunas are dominated by the flaccid, bag like creatures that form much of the Ediacaran Fauna, although there are traces of creatures that could move and graze, and had a rudimentary sense of direction (see: Burrowers: knowing front from back, July 2012 and Something large moved 2 billion years ago). Unsurprisingly, palaeobiologists would like to know when ‘our lot’ arose. One route is via comparative genetics among living animals, using DNA differences and the ‘molecular clock’ approach to estimate the age of evolutionary separation between ‘us’ and ‘them’. But the spread of estimated ages is so broad as to render them almost meaningless. And the better constrained ages of very old trace fossils rely on accepting an assumption that they were, indeed, formed by bilaterians. Yet ingenuity may have revealed an actual early bilaterian from such traces.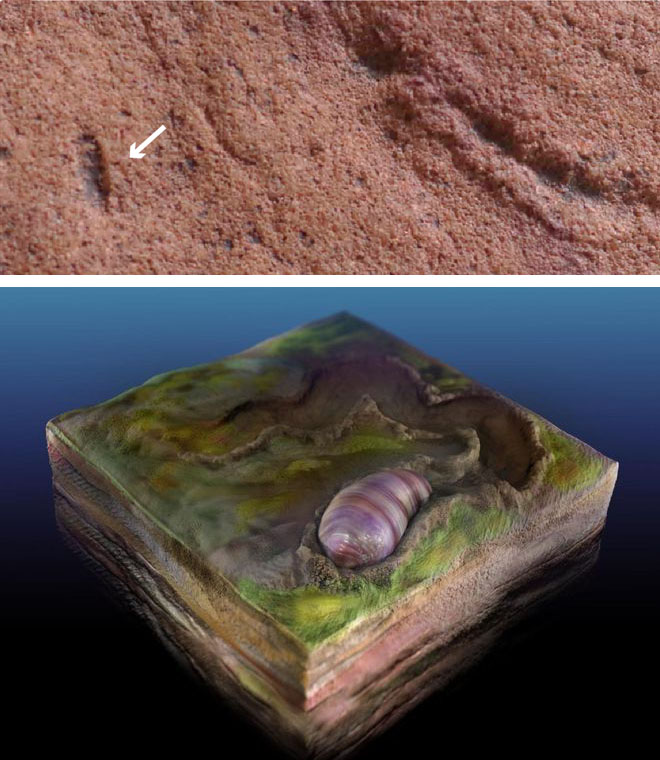
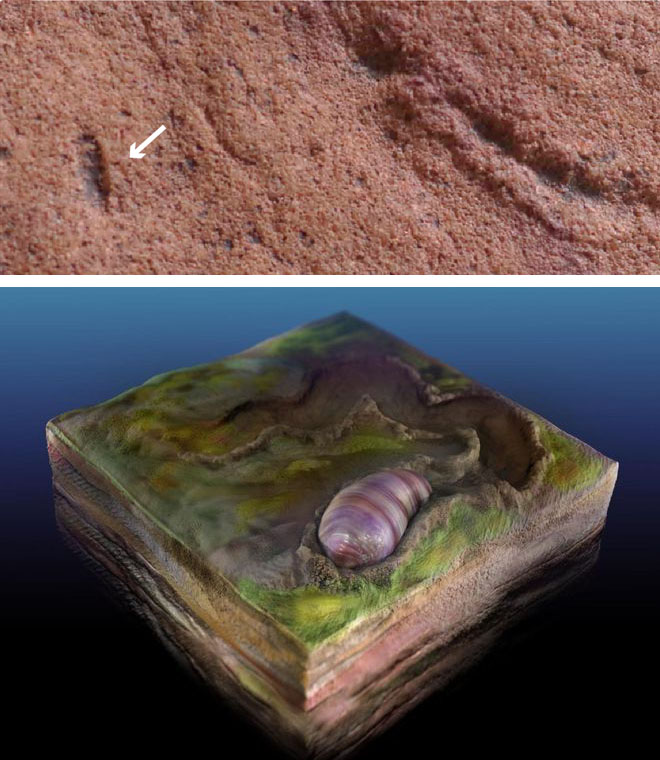
Palaeobiologists from the US and Australia have scoured the famous Ediacara Hills of South Australia for traces of burrowing and signs of the animal that did it (Evans, S.D. et al. 2020. Discovery of the oldest bilaterian from the Ediacaran of South Australia. Proceedings of the National Academy of Sciences, v. 117, online; DOI: 10.1073/pnas.2001045117). One Ediacaran trace fossil, known as Helminthoidichnites is preserved as horizontal trails on the tops and bottoms of thin, discontinuous sand bodies. Luckily, these are sometimes accompanied by elongate ovoids, like large grains of rice. From numerous laser scans of these suspected burrowers, and the traces that they left the authors have reconstructed them as stubby, possibly segmented, worm-like animals that they have called Ikaria wariootia, which may have grazed on algal mats. This name is derived from the local Adnyamathanha people’s word (Ikara or ‘meeting place’) for the locality, a prominent landmark, near Warioota Creek. The age of the sedimentary sequence is between 551 to 560 Ma, and perhaps a little earlier. They could be the earliest-known bilaterians, but the sandy nature of the rocks in which they occur precludes preservation of the necessary detail to be absolutely sure: that would require silt- or. clay-sized granularity
See also: Fossil worm shows us our evolutionary beginnings (BBC, Science and Environment)



