It is always refreshing when physical anthropologists perform experiments as well as pondering on bones. It turns out that examining the bio-mechanics of college baseball players can provide useful clues about where in fossil anatomy to look for signs of potential big-game hunters. Anyone who can hurl a baseball, or one of the smaller but much harder red ones preferred by non-Americans, at speeds exceeding 100 kph could in all likelihood bring down a substantial prey animal with a rock and even more so with a spear. At the heart of an important examination of what our forebears might have done to get a meaty meal (Roach, N.T. et al. 2013. Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo. Nature, v. 498, p. 483-486) is a US-Indian team’s sophisticated study of college baseball players’ throwing action using high-speed video, radar and precise timing techniques.
Matt Kata throwing for the Houston Astros (Photo credit: Wikipedia)
It seems that there are several physiological phases in demon ball throwing: rotation of the torso; rotation flexion and extension of the shoulder; flexion and extension of the elbow; and wrist extension. All of these contribute to acceleration of the ball before release. While the thrower steps forward the arm is cocked so that ligaments, tendons and muscles crossing the shoulder become stretched, thereby storing energy. During the acceleration phase the bend in the elbow is snapped straight adding yet more power. Readers should note the difference between this action and that of a bowler in cricket, where the elbow snap is banned on pain of severe penalty and public humiliation of the bowler who ‘chucks’. Since a fast bowler also adds energy by running into the crease, this is a humanitarian aspect of the Rules of cricket, although several legal West Indian bowlers of the past 40 years are still remembered with terror by their batsmen contemporaries. No such stricture is placed on the baseball pitcher who has no run-up.
These observations focus attention on the structure of shoulder and elbow, yielding a robust means of predicting how fast throwers with different configurations may have thrown objects. Chimpanzees make poor players of ball games, although they will throw the odd stick, but just for aggressive show. The same goes for the earliest hominins for which we have suitable fossil material: australopithecines may occasionally have eaten carrion but they couldn’t throw rocks or spears with enough force to bring down anything and their throwing range would have been pathetic. Not so Homo erectus! They were well equipped in the hurling department and could, were they so inclined, have hunted equally as well as modern humans. Interestingly, earlier hominins had some of the physiological necessities of decent throwing, but not all of them. So it seems that the full combination emerged in the evolution of our own genus around 2 Ma ago,
This is in contrast to a view held by some anthropologists, such as Christopher Boehm of the University of Southern California, that big game hunting using projectile weapons emerged only with anatomically modern humans after 250 ka, and most likely only reached its acme 45 ka ago. That assumption, at least by Boehm, is central to notions of how social activities centred on meat sharing may have helped evolve morals, such as altruism and shame (see Boehm, C. 2012. Moral Origins: The Evolution of Virtue, Altruism and Shame. Basic Books, New York). That H.erectus would have been able to harness sufficient energy to kill at a distance casts doubt on such assertions. Mere foraging does not require throwing-capable physiology, so how it evolved in early humans with neither the inclination nor bodies to at least begin throwing projectiles at potential prey is something that school might consider.
Around 73 thousand years ago a supervolcano in Sumatra erupted on a scale unprecedented in the last 2 million years. It left a 100 by 30 km elliptical caldera now occupied by Lake Toba, and explosively ejected 2800 of magma, about 800 km3 falling as ash as far afield as the Greenland ice cap. Although ice-core records show little if any sign of associated climate change in polar regions, the vast amount of ash and sulfate aerosols blasted into the stratosphere must have had some ‘global winter’ effect. Large areas of South Asia were blanketed by thick beds of ash. Human migration from Africa into Eurasia was probably underway at the time, indeed stone tools are found directly beneath and above the Toba ash in southern India and Malaysia. Some palaeoanthropologists have seen the stresses imposed by the Toba eruption as possible means of reducing the entire human population to a mere few thousand: a genetic ‘bottleneck’ that could have led to rapid evolution among surviving generations that may have shaped changes in human behaviour and culture.
Landsat image of Lake Toba, the largest volcanic crater lake in the world. (credit: Wikipedia)
There is a widening range of views on the climate changes that may have followed Toba. It has even been suggested that global mean surface temperature fell by as much as 10°C (Robock, A. et al. 2009. Did the Toba volcanic eruption of ∼74 ka B.P. produce widespread glaciation?Journal of Geophysical Research: Atmospheres, v. 114, DOI: 10.1029/2008JD011652), although not so far as to produce a worldwide glacial surge but sufficient to devastate vegetation. This bleak look back to a critical point in human affairs resulted from modeling of the effects of a global reflective cloud of ash and sulfate. A later modeling study factored in particle and aerosol sizes (Timmreck, C. et al. 2010. Aerosol size confines climate response to volcanic super-eruptions. Geophysical Research Letters, v. 37, doi:10.1029/2010GL045464) to give a less dramatic, but still severe maximum global cooling due to Toba of ~3.5°C.
The focus has now shifted from modelling to a more direct look at the environmental effects of the Toba super-eruption, preserved in sediments beneath Lake Malawi in southern Africa (Lane, C.S. et al. 2013. Ash from the Toba supereruption in Lake Malawi shows no volcanic winter in East Africa at 75 ka. Proceedings of the National Academy of Science, v. 110, doi/10.1073/pnas.1301474110). The sediments contain a thin ash layer that is very different from those produced by East African Rift volcanism but chemically and texturally similar to the Toba ash from the Indian Ocean and India. The sediments, diatom fossils and chemical biomarkers immediately above the ash show little sign of a significant temperature fall. At most it records a 1.5°C fall, and the authors conclude little chance of a human genetic bottleneck among Africans living at the time.
There is clearly a conflict between results of modeling and real-world climatic data, which is interesting in its own right. But the Malawi findings do not rule out ‘bottlenecks’ resulting from severe stress in South Asia where the ash itself would have severely affected game and vegetation for long enough to face migrating human bands with the prospect of starvation. Obviously, some survived to move on and to leave their tools behind on top of the Toba Ash.
A press release from the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, announces the completion of the most precise genome from a third Neanderthal individual. For the first time it is possible to distinguish copies of the genes inherited by the individual from both parents. The data release coincided with a review of genetic evidence for interbreeding between early Homo sapiens and other species.
Modern human migration out of and within Africa relative to the domains of coeval archaic humans 1 = modern humans 2 = Neanderthals 3 = other archaic humans (credit: Wikipedia)
Liang Bua cave on Flores, Indonesia where fossils of Homo floresiensis were discovered in 2003 (credit: Wikipedia)
When they were first discovered in Liang Bua cave on the Indonesian island of Flores diminutive hominin remains sparked off a heated debate. Part of the reason for dispute was the age of the deposit in which they were found (18 to 850 ka), so young that it indicated possible cohabitation on the island with anatomically modern humans. On the one hand, the finders claimed that they represented a previously unknown hominin species. Other specialists considered that the tiny size (adults no taller than about a metre with brain capacity around that of australopithecines) indicated some congenital dwarfism.
Homo floresiensis skull (credit: Wikipedia)
In the 9 years since the remains came to light, several anatomically features have been cited to support the view of a distinct hominin species: their lack of a chin and different arm and shoulder anatomy, which H. floresiensis shares with H. erectus and H. georgicus. The fossils are associated with simple stone tools and bones of a variety of prey animals that show cut marks and charring, suggesting that cooking was part of these hominins’ lifestyle; despite having small brains they were not unintelligent.
Substantial remains of nine or more individuals have been unearthed so that anatomical detail is almost complete. In 2007 details were published of three well-preserved wrist bones from the original find. They too were sufficiently different from modern and Neanderthal humans to warrant confirmation that H. floresiensis is indeed a distinct hominin species. Further work on wrist bones from other individuals has now more or less put the seal on this identity (Orr, C.M. et al. 2013. New wrist bones of Homo floresiensis from Liang Bua (Flores, Indonesia). Journal of Human Evolution, v. 64, p. 109-129), the authors concluding that ‘The pattern of morphology … supports H. floresiensis as a valid taxon and refutes the hypothesis that these specimens represent modern humans with some kind of pathology or growth disturbance’. They take matters further by suggesting that their lineage was established before divergence of modern humans and Neanderthals. As with the shoulder morphology that of their wrists would have somewhat hindered tool-making dexterity, but nonetheless they did make tools.
Scientific American is renowned for its eminently readable reviews of both emerging and perennial topics. Its February 2013 issue takes on one that is guaranteed to run and run; the evolutionary course that produced us (Harman, K. 2013. Shattered ancestry. Scientific American, v. 308 (February 2013), p. 36-43). Since its launch Earth Pages has covered much of the new science in the field but did not anticipate the depth of the stir towards which it has led.
For a decade it has become increasingly clear that anatomically modern humans are unique in one respect: they are the first species in perhaps 4 million years to be the sole extant member of the cladistic tribe Hominini. As recently as 30 ka Homo sapiens shared the planet with Neanderthals, Denisovans, H. erectus and H. floresiensis. At the time the genus Homo emerged around 2.0-2.5 Ma ago there were at least four other fossil groups that shared the major characteristic of upright gait, all australopithecines in ‘robust’ and ‘gracile’ guises.
As time goes by there will likely be more fossil discoveries that show important anatomical signs of other novel evolutionary divergence, which therefore warrant new species. Pliocene-Pleistocene time is becoming crowded, and the more diversity in its fossil record the less likely it is that some clear evolutionary pathways can be devised to explain just what was going on. Katherine Harmon of Scientific American’s editorial team touches on the thorny issues of upright walking and gait, tree climbing, precise use of the fingers and thumb, and brain size that are raised by 22 species; 2 living and 20 extinct.
Genetics clearly indicates that our nearest living relatives belong to two species in the genus Pan(chimpanzees and bonobos). It has been generally assumed that the common ancestor of this extant kinship some 8 Ma back was chimp-like, and that evolutionary divergence from its habits and anatomy produced the growing ‘bramble patch’ of hominin evolution. That assumption is based on the principle of parsimony, i.e. the simplest view of the evidence – what there is now and fragments from the past eight million years. The trouble is there is a dearth of fossils that can be said to be en route to chimps in some way.
In fact today’s chimps and bonobos are more or less restricted to clambering in tropical forest habitats, for which they are well-adapted. Maybe they are the survivors of evolutionary vagaries just as complex as those leading to us. For one thing, almost embarrassingly, their brain size is substantially larger than those of quite a few fossil hominins; and why not? How they behave socially may possibly have arisen as part of their specialisation too, of which more shortly. Our big difference from them is being supreme generalists, as well as consciousness.
All the fossils classed as hominins show some signs of being able to walk upright, classically the forward position of the foramen magnum at the base of the skull where it joins to the backbone, but in some cases merely the geometry of the hip joint to the pelvis for that is all that has been found. Yet that anatomical likelihood glosses over the vital detail of the actual gait – heel-to-toe like us (Australopithecus afarensis), on the outside edge of the foot akin to chimps (Ardepithecus ramidus) or differently again but possible as efficient as us (Au. sediba). Then there is the matter of arboreal abilities: chimps are masters despite their bulk, but every hominin whose foot bones have been found shows some evidence of grasping with the big toe. Indeed humans are pretty nimble climbers but do not brachiate from branch to branch.
As regards the hands, an interesting point is that while chimpish knuckle walking is not seen in fossils, Ardipithecus probably could walk on all fours with hands flat on the ground but had fingers quite capable of precise manipulation, an ability shown spectacularly well by 2 Ma old Au. sediba. Upright walking may have evolved more than once, and it is even possible that chimps evolved specifically for climbing in forestlands, their highly adapted grasping hands only capable of knuckle walking on the ground.
Oreopithecus bambolii from the Upper Miocene of northern Italy(credit: Wikipedia)
The late-Miocene of Africa – the likely time range for the Pan-Homo common ancestor – is a fossil desert as regards primates. Yet its Italian equivalent has yielded a fascinating and well-preserved creature; Oreopithecus bambolii has skeletal features compatible with an upright posture and bipedal locomotion. Until the African Miocene yields something more appropriate, Oreopithecus is a candidate for a common ancestor, and interesting in another respect. Its dentition does not include prominent canine teeth that in the predominantly vegetarian, though occasionally carnivorous, Pan species serve well in their aggression-based, hierarchical social systems, as they do in the even more spectacular baboons.
Christopher Boehm, primate behaviouralist cum anthropologist, in his recent book Moral Origins (2012 Basic Books, ISBN-13: 978-0465020485) uses the principle of parsimony to reconstruct the social system of the Miocene Pan-Homo common ancestor from those of chimps and surviving human hunter-gatherers. His thesis is that it was centred on the hierarchical dominance of ‘alpha’ males, as is that of chimps. Prolonged social selection in hominin evolution largely tempered such a ‘Big Man’ tendency through a variety of strategies directed by majorities. Social punishments, including capital punishment, evolved to combat free-loading, theft and individual dominance in favour of cooperative egalitarianism. Such measures developed increasingly conscious self-suppression of such traits that eventually manifested themselves as what we now regard as human morals. Boehm considers that this psychological trend in evolution accelerated once Homo sapiens began hunting of large prey animals that added substantially to diet.
There is a major problem for this view: like Oreopithecus every well-preserved hominin species, even the earliest Sahelanthropus tchadensis, do not have prominent canines irrespective of whether they show evidence of at least partial meat-eating or pure vegetarianism. For some species with many fossil members, such as Au. afarensis, there are signs ofsexual dimorphism – larger males than females – but that does not necessarily signify hierarchical social behaviour. With the appearance of H. erectus that difference wanes to the present slight differences between modern male and female humans.
Agressive male humans, note gumshields (credit: John_Scone via Flickr)
If it is valid – and who knows? – for morphology to give clues to social behaviour, then it is equally likely that the beginnings of the hominin evolutionary thicket may well have involved a trend in social behaviour towards cooperative action; 8 million years ago. For generally small, gracile creatures with habits no more threatening to the big predators of the African savannahs that that of the porcupine, there would have been a powerful selection pressure towards a united front. Of course, in the last ten thousand years since the shift to economic strategies based on storable surpluses and their expropriation, hierarchical social systems with violence at their heart emerged among modern humans. Judging by the body shapes and dentition of extant ‘alphas’, as in capital’s boardrooms and among the frontbenchers at Westminster, anthropology clearly is in need of some refinement…
Landsat image of the Lake Toba caldera, Sumatra (credit: Wikipedia)
The largest volcanic catastrophe during the evolution of humans formed the huge caldera at Lake Toba near the Equator in Sumatra about 70 thousand years ago. Explosive action erupted 2800 cubic kilometres of magma, of which 800 km3 was deposited as thick ash across most of South Asia and the northern Indian Ocean. Sulfates derived from the gas emissions by Toba form clear ‘spikes’ in ice cores from both Greenland and Antarctica. Its effects were global through the mixing of sulfate aerosols in the stratosphere of both hemispheres, encouraged by its position close to the Equator. By reflecting incoming solar energy the aerosols resulted in a century-long 10°C fall in temperature over the Greenland ice cap. Such global cooling almost certainly affected anatomically modern humans, but it is possible that in South Asia Toba had an even more devastating effect.
The Toba ash at the Jwalapuram excavations in South India(Photo credit: Sanjay P. K. via Flickr)
At several sites in the Indian state of Tamil Nadu and in Malaysia Toba ash has buried artifacts that arguably may have been made by the earliest modern emigrants from Africa. Immediately above the ash are yet more tools that suggest humans did survive the eruption. Palaeoanthropologists have argued that the stress of Toba’s environmental effects on all hominins living at the time may have resulted in population crashes from which only the fittest individuals emerged. Major evolutionary changes have been ascribed to ‘bottlenecks’ of that kind to result in changes in human behaviour detectable from the archaeological record, such as the creation of completely new kinds of tools, art and language. However, recent finds in Africa suggest that many such shifts are much older than Toba.
Perhaps Toba’s greatest contribution to palaeoanthropology is that it is an easily recognised event in the geological record, but compared with its sulfate spike in the Greenland ice core at ~71 ka the existing radiometric dates have uncertainties of several thousand years. Using the latest 40Ar/39Ar dating methods on fresh crystals of sanidine (volcanic K-feldspar) from new excavations in Malaysia these uncertainties have been reduced significantly (Storey, M. et al. 2012. Astronomically calibrated 40Ar/39Ar age for the Toba supereruption and global synchronization of late Quaternary records. Proceedings of the National Academy of Sciences, v. 109, p. 19684-18688 ). The sulfate peak and the ash can now be attributed to an age of 73.88 ± 0.32 ka; better than a golden spike in Late Pleistocene stratigraphy. The ice-cores have a check on chronology just beyond the limit of counting annual layering, as do ocean sediment cores for a time older than 14C can ever achieve. Toba now links too with events recorded by the precise U-Th series dating of cave deposits
If Ignatius Loyola been a child of the late 20th century, it is quite likely that he would have chosen palaeoanthropology as a career rather than theology, seeing as he was so predisposed to casuistry. When I innocently asked a vertebrate palaeontologist who specialized in the Pliocene and Pleistocene Epochs why it was that students of hominins were so prone to controversy, his answer was revealing: ‘They don’t have many fossils’. One place where there are lots of hominin fossils, in fact the largest known sample of them, is the Atapuerca cavern in northern Spain. At the deepest level of the cave system there is a veritable charnel house containing the remains of at least 28 individuals. Because there are bones from all parts of the human anatomy, some have suggested that the cache is one of deliberate burial, but there is a disturbing dearth of the smaller bones of feet and hands. Consequently, other voices claim that the bodies were washed in by floods, losing extremities en route – though that view would be easily tested using other signs of trauma on large bones. Yet that is a minor quibble compared with one that is developing around the age of the boneyard and the taxonomy of the cadavers in it (http://www.guardian.co.uk/science/2012/jun/10/fossil-dating-row-sima-huesos-spain).
Head of Homo heidelbergensis , Senckenberg Museum, Frankfurt am Main, Germany (Photo credit: Wikipedia)
The Spanish team responsible for the evolutionary wealth in the entire Atapuerca cave complex, which ranges from almost a million years ago to recent times, assigned the Sima de los Huesos (Pit of Bones) fossils to Homo heidelbergensis. In fact about 90% of all H. heidelbergensis remains are from Atapuerca, so any anatomical dispute over these specimens is a threat to the status of the species itself. One leading authority who does dispute this assignment is Chris Stringer of the UK Natural History Museum, who claims that many of the heads have teeth and jaws with shapes that fall within the range of Neanderthals – supposedly descended from H. heidelbergensis. The age of the deposit is the focus of debate. Were it to be around 400 ka or younger, as early attempts at dating suggested, then the fossils might well be those of Neanderthals for that is early in the range of that species as determined by ‘molecular-clock’ studies of Neanderthal DNA. However, the material most likely to yield a good radiometric age is carbonate speleothem, the stuff of stalactites and stalagmites though more commonly a matrix that binds together old cave detritus. The fossils are undoubtedly far older than the maximum age that can be achieved using the well known radiocarbon method (<60 ka), but speleothem lends itself to a precise dating technique based on the decay series of uranium isotopes. In the case of Sima de los Huesos, the fossils lie in a clay breccia overlain by a layer of speleothem, which has yielded a U-series age of around 600 Ma (Bischoff, J.L. et al. 2007. High-resolution U-series dates from the Sima de los Huesos hominids yields 600 kyrs: implications for the evolution of the early Neanderthal lineage. Journal of Archaeological Science, v. 34, p. 763-770).
The ‘bone breccia’ in Sima de los Huesos, Atapuerca caverns Spain (from Bischoff, J.L. et al. 2007)Neanderthal head from Israel (Wikipedia)
Stringer argues that the hominins’ anatomy is so like that of Neanderthals that, somehow, the radiometric age must be wrong – i.e. “too old” – perhaps because the speleothem is in fact from a 600 ka block that fell onto the fossils after they had accumulated. His view is that they are Neanderthals descended from H. heidelbergensis living in the earlier Pleistocene and which was the common ancestor of both Neanderthals and anatomically modern humans. Bischoff et al. consider the Sima de los Huesos hominids to be ‘at the very beginnings of the Neanderthal evolutionary lineage’, which seems to me to be a reasonable deduction from both stratigraphic and anatomical data. To demand that they must be at least 200 ka younger, apparently on the basis of an estimate of Neanderthal origination from DNA data seems less reasonable. The appearance of Stringer’s detailed arguments in Evolutionary Anthropology (v. 21(3)) is eagerly awaited, following the Observer’s take on his position.
Another area in which controversy is brewing – and has been for decades – is that of the origin of human artistic culture. One of the gem-boxes of early art is the Geissenclösterle (monastery of the goats) cavern in southern Germany, in which have been found various figurines made of bird bone and ivory, including a celebrated lion-man theriomorph, highly exaggerated female figures, flutes and beads. They belong to the Aurignacian culture brought by the earliest anatomically modern Europeans who diffused westwards along the Danube from the near-East as early as 45 ka ago. The layer containing the artifacts was originally dated at about 35 ka, but new radiocarbon techniques have been tried on bone with cut marks, among other materials (Higham, T. et al. 2012. Testing models for the beginnings of the Aurignacian and the advent of art and music: the radiocarbon chronology of Geissenclösterle. Journal of Human Evolution, v. 62, p. 664-676 doi:10.1016/j.jhevol.2012.03.003) and found to yield a much older age of 42.5 ka, close to the oldest European date for modern human occupation 43-45 ka for the stratigraphically older Uluzzian tool industry.
Lion-man sculpture from Geissenclösterle ( J. Duckek Wikipedia)
The date is also considerably earlier than the demise of the Neanderthals and raises the issue of modern-Neanderthal contacts. Indeed the layer below that assigned to Aurignacian contains tools made by Neanderthals, whose age is statistically indistinguishable from the later occupation level. The Chatelperronian tool industry, which closely resembles the Aurignacian but is ascribed to Neanderthals, is supposed to be around 40 ka old, but the advanced radiocarbon technique that yielded much older ages for Geissenclösterle apparently has not yet been deployed on this culture. On the basis of limited age data, it does seem likely that Neanderthals adopted the new technology after they encountered it. The Aurignacian artistic products are vastly more advanced than any found at older sites in Africa.
Aurignacian female figurine from near Geissenclösterle..(Silosarg: Wikipedia)
In the context of the debate about modern human and Neanderthal cognitive abilities, which suggests the former were altogether smarter and more creative, there is an unvoiced or at least unheeded argument. Whether or not Neanderthals originated artifacts that were ‘modern’ for their time or copied them is not as important as the fact that this group, previously isolated for up to 400 millennia, were able to appreciate and learn these novelties. That is much the same as people living today, in Australia for instance, a couple of generations from hunter-gatherer origins, working on production lines, piloting aircraft, social networking and creating world-class abstract art. What did they, and the Aurignacians, produce from other materials that have not survived decay; ditto for any pre-45 ka humans? Another point rarely raised, but surely valid, is that previous people may not have felt any need to produce art in forms that survive for tens or hundreds of millennia. Forty-odd thousand years ago, climate was undergoing rapid ups and downs of temperature and humidity in the run-up to the last glacial maximum. Conditions at mid-latitudes would have been much more changeable than those of the tropics. Both anatomically modern humans and Neanderthals faced the same attendant ecological changes, and as co-occupants of southern Europe they faced each other as rivals for available resources. Finally, Aurignacians hailed from the east, also Neanderthal territory and severely affected by rapid climate change from around 80 ka; so did they bring with them a culture formed elsewhere? Europe concentrates palaeoanthropologists and their endeavours, while much of the planet to which humans diffused from Africa – and Africa itself – are grossly under-investigated by comparison: ideas will undoubtedly change drastically as these areas get the attention they deserve.
Controversy is not a problem. Indeed, with imperfect, inadequate or ambiguous data it is unavoidable, and heated disputes spur the search for more information that can help resolve ideas or change them. What cannot be sidestepped is the potential for havoc that may arise with new and improved methods. In both cases outlined here radiometric dates have thrown the proverbial spanner into the works. The method used in the Geissenclösterle cavern was designed to remove younger contaminating material from samples for radiocarbon dating and inevitably tends to push 14C dates further back in time. By removing a source of inaccuracy it highlights the inadequacies of dates obtained by earlier approaches on which a great deal of current archaeological thinking relies. Just how much younger contamination is present in a sample only emerges after the improved dating: it may be absent but an be substantial. So, until materials dated by earlier radiocarbon methods are re-run using the new approach neither their absolute ages nor their relative sequence in time can be considered reliable.
Art on the walls of Altamira Cave, northern Spoain, including both older abstract works and younger figurative depictions of prey animals (Photo credit: Wikipedia)
Results from just such an advance in radiometric dating of cave deposits in northern Spain will really cause a stir, when they sink in (Pike, A.W.G. and 10 others 2012. U-series dating of Paleolithic art in 11 caves in Spain. Science, v. 336, p. 1409-1413). The U-series method used at the University of Bristol by the joint British-Spanish collaborators dates calcite deposits on painted cave walls, including those at the famous Altamira site. This ‘flowstone’ may underlie artwork or may have grown over it after its completion, giving maximum or minimum ages for the painting, respectively. If a work has flowstone underneath and as a coating, dating potentially ‘brackets’ a possible age range. The superb figurative depictions of various prey animals, such as bison in Altamira cave, turn out to have been painted at around 18 ka, during the last glacial maximum. However a lot of the art there is abstract, such as hands picked out by red pigment presumably sprayed onto the wall from the artist’s mouth, various stippled discs and dots. Many of the abstracts are beneath flowstone that is around twice as old as the more familiar objects and range in age from 34 to 41 ka, thereby being close in time with the Geissenclösterle materials. Like them, their ages may coincide with the arrival of the earliest anatomically modern Europeans, but they are also towards the end of the period when Neanderthals were still present in much of Europe, including northern Spain. It cannot be ruled out therefore that the earliest paintings were Neanderthal symbolic art.
Related articles
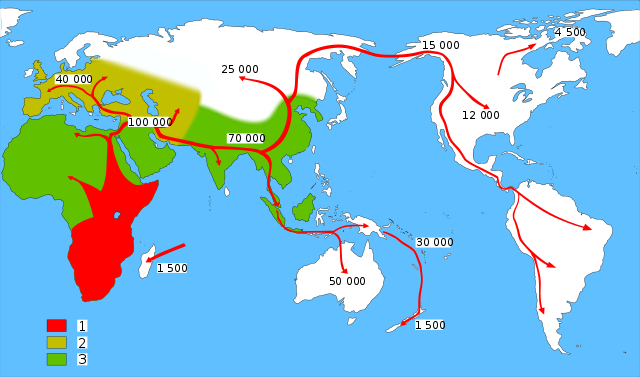
The 3 May 2012 issue of Nature contained a broad review of human migration (Peopling the Planet p. 23-35) including a summary of unpublished precision dating (Callaway, E. Date with history. p. 27-29)
Balter, M. 2012. New light on revolutions that weren’t. Science, v. 336. p. 530-531







")








, Sencke...")






