Sediments built up on lake beds are a fruitful source of proxy data for all kinds of time series – mainly climatic and ecological. Pollen, other organic remains, various stable isotopes, and a range of organic geochemical data calibrated to time using magnetostratigraphy, C-14 dating and astronomical ‘pacemakers’. Suddenly there is another proxy: cannabinol, the metabolite of tetrahydrocannibinol the principal psychoactive component of marijuana (Lavrieux, M. et al. 2013. Sedimentary cannabinol tracks the history of hemp retting. Geology, v. 41, p. 751-754). The compound is detectable at the parts per billion level thanks to advances in monitoring the use of drugs, particularly in sports persons – it ends up in the urine of users. So the paper by a team of French Earth scientists has a somewhat irresistible draw, the more so from the opening sentence of its abstract, ‘Hemp (Cannabis sp.) has been a fundamental plant for the development of human societies’. Indeed it has, for the earliest records date back to the Neolithic in China, perhaps back to 12 ka ago.
Cultivation of hemp for fibre and grain in France. (credit: Wikipedia)
But then all becomes clear: they speak of hemp fibres used in rope and some textiles, and the climatic adaptability of the plant that has ensured its spread from Equator to north of the Arctic Circle and lesser southern latitudes. But there is an element of tongue-in-cheek, or at least so it seemed to me, as the objective of their research is to chart to emergence and rise of rope making in Central France. Freeing the useful fibres from Cannabis stems requires the plant to be soaked and subject to microbial action that breaks down soft tissue, know as retting that is also used in flax and coir production. The resin breaks down to cannabinol, which is therefore a perfect proxy for Hemp retting.
Lac d’Aydat is geologically famous as it formed when a lava flow from one of the puys of the Massif Central blocked a valley and became a dam. It figured in the pioneering volcanological research of English geologist George Julius Poulett Scrope. Its new place in science rests on Lavrieux et al.’s chronologically calibrated time series for retting from the lake’s muds. Hemp pollen in the section betrays the start of Cannabis cultivation in the Auvergne between 500-650 AD, but hemp retting in the lake is marked by a cannabinol spike in the 13th century and increases in pollen. It fell-off sharply in the late 19th century, probably as a result of being outcompeted by more easily processed cotton.
Almost 7 centuries of Cannabis processing in central France actually took a toll as cannabinol is toxic to fish and cattle. Despite a 1669 Royal Ordinance against hemp retting in French rivers it continued unchecked in Lac d’Aydat, but more likely than secret retting tucked away in a remote corner of France it stemmed from the ordinance being widely flouted. That it ended with the rise of cotton is not so convincing as hemp is still a staple in rope manufacture, and when the US entered World War II large tracts of land were placed under Cannabis to produce naval cordage; the reason why it still grows wild in abundance across many States. There is plenty of evidence, including this, that use of Cannabis for cordage came rather late, and plenty in support of its cultivation and wide spread before the Iron Age for ‘relaxation’.
It is always refreshing when physical anthropologists perform experiments as well as pondering on bones. It turns out that examining the bio-mechanics of college baseball players can provide useful clues about where in fossil anatomy to look for signs of potential big-game hunters. Anyone who can hurl a baseball, or one of the smaller but much harder red ones preferred by non-Americans, at speeds exceeding 100 kph could in all likelihood bring down a substantial prey animal with a rock and even more so with a spear. At the heart of an important examination of what our forebears might have done to get a meaty meal (Roach, N.T. et al. 2013. Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo. Nature, v. 498, p. 483-486) is a US-Indian team’s sophisticated study of college baseball players’ throwing action using high-speed video, radar and precise timing techniques.
Matt Kata throwing for the Houston Astros (Photo credit: Wikipedia)
It seems that there are several physiological phases in demon ball throwing: rotation of the torso; rotation flexion and extension of the shoulder; flexion and extension of the elbow; and wrist extension. All of these contribute to acceleration of the ball before release. While the thrower steps forward the arm is cocked so that ligaments, tendons and muscles crossing the shoulder become stretched, thereby storing energy. During the acceleration phase the bend in the elbow is snapped straight adding yet more power. Readers should note the difference between this action and that of a bowler in cricket, where the elbow snap is banned on pain of severe penalty and public humiliation of the bowler who ‘chucks’. Since a fast bowler also adds energy by running into the crease, this is a humanitarian aspect of the Rules of cricket, although several legal West Indian bowlers of the past 40 years are still remembered with terror by their batsmen contemporaries. No such stricture is placed on the baseball pitcher who has no run-up.
These observations focus attention on the structure of shoulder and elbow, yielding a robust means of predicting how fast throwers with different configurations may have thrown objects. Chimpanzees make poor players of ball games, although they will throw the odd stick, but just for aggressive show. The same goes for the earliest hominins for which we have suitable fossil material: australopithecines may occasionally have eaten carrion but they couldn’t throw rocks or spears with enough force to bring down anything and their throwing range would have been pathetic. Not so Homo erectus! They were well equipped in the hurling department and could, were they so inclined, have hunted equally as well as modern humans. Interestingly, earlier hominins had some of the physiological necessities of decent throwing, but not all of them. So it seems that the full combination emerged in the evolution of our own genus around 2 Ma ago,
This is in contrast to a view held by some anthropologists, such as Christopher Boehm of the University of Southern California, that big game hunting using projectile weapons emerged only with anatomically modern humans after 250 ka, and most likely only reached its acme 45 ka ago. That assumption, at least by Boehm, is central to notions of how social activities centred on meat sharing may have helped evolve morals, such as altruism and shame (see Boehm, C. 2012. Moral Origins: The Evolution of Virtue, Altruism and Shame. Basic Books, New York). That H.erectus would have been able to harness sufficient energy to kill at a distance casts doubt on such assertions. Mere foraging does not require throwing-capable physiology, so how it evolved in early humans with neither the inclination nor bodies to at least begin throwing projectiles at potential prey is something that school might consider.
Around 73 thousand years ago a supervolcano in Sumatra erupted on a scale unprecedented in the last 2 million years. It left a 100 by 30 km elliptical caldera now occupied by Lake Toba, and explosively ejected 2800 of magma, about 800 km3 falling as ash as far afield as the Greenland ice cap. Although ice-core records show little if any sign of associated climate change in polar regions, the vast amount of ash and sulfate aerosols blasted into the stratosphere must have had some ‘global winter’ effect. Large areas of South Asia were blanketed by thick beds of ash. Human migration from Africa into Eurasia was probably underway at the time, indeed stone tools are found directly beneath and above the Toba ash in southern India and Malaysia. Some palaeoanthropologists have seen the stresses imposed by the Toba eruption as possible means of reducing the entire human population to a mere few thousand: a genetic ‘bottleneck’ that could have led to rapid evolution among surviving generations that may have shaped changes in human behaviour and culture.
Landsat image of Lake Toba, the largest volcanic crater lake in the world. (credit: Wikipedia)
There is a widening range of views on the climate changes that may have followed Toba. It has even been suggested that global mean surface temperature fell by as much as 10°C (Robock, A. et al. 2009. Did the Toba volcanic eruption of ∼74 ka B.P. produce widespread glaciation?Journal of Geophysical Research: Atmospheres, v. 114, DOI: 10.1029/2008JD011652), although not so far as to produce a worldwide glacial surge but sufficient to devastate vegetation. This bleak look back to a critical point in human affairs resulted from modeling of the effects of a global reflective cloud of ash and sulfate. A later modeling study factored in particle and aerosol sizes (Timmreck, C. et al. 2010. Aerosol size confines climate response to volcanic super-eruptions. Geophysical Research Letters, v. 37, doi:10.1029/2010GL045464) to give a less dramatic, but still severe maximum global cooling due to Toba of ~3.5°C.
The focus has now shifted from modelling to a more direct look at the environmental effects of the Toba super-eruption, preserved in sediments beneath Lake Malawi in southern Africa (Lane, C.S. et al. 2013. Ash from the Toba supereruption in Lake Malawi shows no volcanic winter in East Africa at 75 ka. Proceedings of the National Academy of Science, v. 110, doi/10.1073/pnas.1301474110). The sediments contain a thin ash layer that is very different from those produced by East African Rift volcanism but chemically and texturally similar to the Toba ash from the Indian Ocean and India. The sediments, diatom fossils and chemical biomarkers immediately above the ash show little sign of a significant temperature fall. At most it records a 1.5°C fall, and the authors conclude little chance of a human genetic bottleneck among Africans living at the time.
There is clearly a conflict between results of modeling and real-world climatic data, which is interesting in its own right. But the Malawi findings do not rule out ‘bottlenecks’ resulting from severe stress in South Asia where the ash itself would have severely affected game and vegetation for long enough to face migrating human bands with the prospect of starvation. Obviously, some survived to move on and to leave their tools behind on top of the Toba Ash.
Australopithecus sediba 1 (MH1) left, Au. afarensis( AL 288-Lucy) centre and Au. sediba 2 (MH2) right. (credit: L. R. Berger, University of the Witwatersrand, via Wikipedia)
The remarkable find of two well-preserved skeletons of a 2 Ma hominin in a South African cave in 2008 and publication of their preliminary analysis in 2011 seemed set to shake up human origins research. There was a more or less complete hand – indeed an entire arm and shoulder – a lower leg with ankle bones, a near-complete head and lots more besides. Most was from one female individual, but significant bits from two others that allowed a well-supported reconstruction of the new species Au. sediba. The discoverer, Lee Berger of The University of the Witwatersrand, South Africa (well he initiated the dig, but his young son found the first critical material) is so excited 5 years on that he uses hip-hop phraseology, she ‘got swag’, presumably assuming that means pretty cool (Gibbons, A. 2013. A human smile and funny walk for Australopithecus sediba. Science, v. 340, p. 132-133), but on the street there are other meanings and attitudes towards the phrase and unwary use is not advised.
More details now have emerged in a special issue of Science introduced by Berger in less fulsome language (Berger, L.R. 2013. The mosaic nature of Australopithecus sediba. Science, v. 340, p. 163). As the title suggests, the surprise lies in almost every critical part of the species. Although the spine shows curvature (lordosis) needed for an animal evolved from a quadruped to bipedality in order to balance when upright, the ankle bone is unlike the flat-based human one, being pointed as is that of chimpanzees. As a result walking would have involved an unusual and perhaps unsteady gait; the individuals did fall over into a death pit and one commentator thought the gait might have seemed ‘provocative’. An unusual knee bone is thought to be an evolved countermeasure to such exaggerated mincing. Despite the very human-like hand, extremely long arms and shoulders remarkably like those of the favoured jacket of a star of the BBC series The Dragons’Den point to habitual clambering in trees. Authors of a report on dentition suggest a close similarity to that of the Au. africanus, living at the same time and also found in the same system of fossil-rich caves north-west of Johannesburg, South Africa. Controversially, the tooth team suggests a closer similarity of both to early Homo species than to earlier australopithecines in East Africa, which would shift the focus of human origins to southern Africa. Counter to that view is a find of 400 ka-older, putative human remains in Ethiopia. Yet they take the form of a lower jaw that resembles that of Au. sediba.
The emerging, more detailed picture is not tidy, as suspected from early examination of the Malapa hominins. One thing is for sure, the South African caves are being swarmed over, which paid dividends in 2011 just 15 km from the Malapa cave with another embarrassment of riches at Sterkfonein in the form of abundant foot bones of a currently un-named species of roughly the same age. Things are beginning to take on an element of national pride, with ‘The Birthplace’ at stake: Kenya, Tanzania, Ethiopia or South Africa?
A press release from the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, announces the completion of the most precise genome from a third Neanderthal individual. For the first time it is possible to distinguish copies of the genes inherited by the individual from both parents. The data release coincided with a review of genetic evidence for interbreeding between early Homo sapiens and other species.
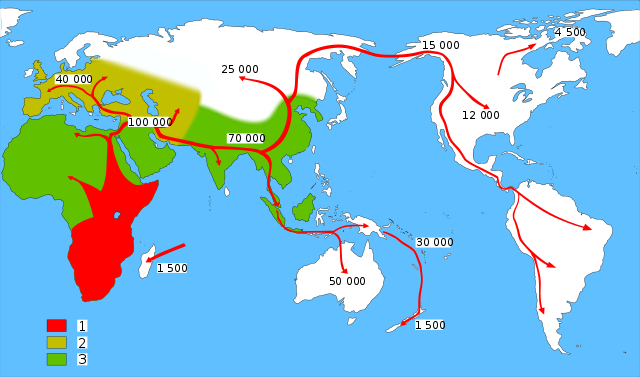
Modern human migration out of and within Africa relative to the domains of coeval archaic humans 1 = modern humans 2 = Neanderthals 3 = other archaic humans (credit: Wikipedia)
Liang Bua cave on Flores, Indonesia where fossils of Homo floresiensis were discovered in 2003 (credit: Wikipedia)
When they were first discovered in Liang Bua cave on the Indonesian island of Flores diminutive hominin remains sparked off a heated debate. Part of the reason for dispute was the age of the deposit in which they were found (18 to 850 ka), so young that it indicated possible cohabitation on the island with anatomically modern humans. On the one hand, the finders claimed that they represented a previously unknown hominin species. Other specialists considered that the tiny size (adults no taller than about a metre with brain capacity around that of australopithecines) indicated some congenital dwarfism.
Homo floresiensis skull (credit: Wikipedia)
In the 9 years since the remains came to light, several anatomically features have been cited to support the view of a distinct hominin species: their lack of a chin and different arm and shoulder anatomy, which H. floresiensis shares with H. erectus and H. georgicus. The fossils are associated with simple stone tools and bones of a variety of prey animals that show cut marks and charring, suggesting that cooking was part of these hominins’ lifestyle; despite having small brains they were not unintelligent.
Substantial remains of nine or more individuals have been unearthed so that anatomical detail is almost complete. In 2007 details were published of three well-preserved wrist bones from the original find. They too were sufficiently different from modern and Neanderthal humans to warrant confirmation that H. floresiensis is indeed a distinct hominin species. Further work on wrist bones from other individuals has now more or less put the seal on this identity (Orr, C.M. et al. 2013. New wrist bones of Homo floresiensis from Liang Bua (Flores, Indonesia). Journal of Human Evolution, v. 64, p. 109-129), the authors concluding that ‘The pattern of morphology … supports H. floresiensis as a valid taxon and refutes the hypothesis that these specimens represent modern humans with some kind of pathology or growth disturbance’. They take matters further by suggesting that their lineage was established before divergence of modern humans and Neanderthals. As with the shoulder morphology that of their wrists would have somewhat hindered tool-making dexterity, but nonetheless they did make tools.
Scientific American is renowned for its eminently readable reviews of both emerging and perennial topics. Its February 2013 issue takes on one that is guaranteed to run and run; the evolutionary course that produced us (Harman, K. 2013. Shattered ancestry. Scientific American, v. 308 (February 2013), p. 36-43). Since its launch Earth Pages has covered much of the new science in the field but did not anticipate the depth of the stir towards which it has led.
For a decade it has become increasingly clear that anatomically modern humans are unique in one respect: they are the first species in perhaps 4 million years to be the sole extant member of the cladistic tribe Hominini. As recently as 30 ka Homo sapiens shared the planet with Neanderthals, Denisovans, H. erectus and H. floresiensis. At the time the genus Homo emerged around 2.0-2.5 Ma ago there were at least four other fossil groups that shared the major characteristic of upright gait, all australopithecines in ‘robust’ and ‘gracile’ guises.
As time goes by there will likely be more fossil discoveries that show important anatomical signs of other novel evolutionary divergence, which therefore warrant new species. Pliocene-Pleistocene time is becoming crowded, and the more diversity in its fossil record the less likely it is that some clear evolutionary pathways can be devised to explain just what was going on. Katherine Harmon of Scientific American’s editorial team touches on the thorny issues of upright walking and gait, tree climbing, precise use of the fingers and thumb, and brain size that are raised by 22 species; 2 living and 20 extinct.
Genetics clearly indicates that our nearest living relatives belong to two species in the genus Pan(chimpanzees and bonobos). It has been generally assumed that the common ancestor of this extant kinship some 8 Ma back was chimp-like, and that evolutionary divergence from its habits and anatomy produced the growing ‘bramble patch’ of hominin evolution. That assumption is based on the principle of parsimony, i.e. the simplest view of the evidence – what there is now and fragments from the past eight million years. The trouble is there is a dearth of fossils that can be said to be en route to chimps in some way.
In fact today’s chimps and bonobos are more or less restricted to clambering in tropical forest habitats, for which they are well-adapted. Maybe they are the survivors of evolutionary vagaries just as complex as those leading to us. For one thing, almost embarrassingly, their brain size is substantially larger than those of quite a few fossil hominins; and why not? How they behave socially may possibly have arisen as part of their specialisation too, of which more shortly. Our big difference from them is being supreme generalists, as well as consciousness.
All the fossils classed as hominins show some signs of being able to walk upright, classically the forward position of the foramen magnum at the base of the skull where it joins to the backbone, but in some cases merely the geometry of the hip joint to the pelvis for that is all that has been found. Yet that anatomical likelihood glosses over the vital detail of the actual gait – heel-to-toe like us (Australopithecus afarensis), on the outside edge of the foot akin to chimps (Ardepithecus ramidus) or differently again but possible as efficient as us (Au. sediba). Then there is the matter of arboreal abilities: chimps are masters despite their bulk, but every hominin whose foot bones have been found shows some evidence of grasping with the big toe. Indeed humans are pretty nimble climbers but do not brachiate from branch to branch.
As regards the hands, an interesting point is that while chimpish knuckle walking is not seen in fossils, Ardipithecus probably could walk on all fours with hands flat on the ground but had fingers quite capable of precise manipulation, an ability shown spectacularly well by 2 Ma old Au. sediba. Upright walking may have evolved more than once, and it is even possible that chimps evolved specifically for climbing in forestlands, their highly adapted grasping hands only capable of knuckle walking on the ground.
Oreopithecus bambolii from the Upper Miocene of northern Italy(credit: Wikipedia)
The late-Miocene of Africa – the likely time range for the Pan-Homo common ancestor – is a fossil desert as regards primates. Yet its Italian equivalent has yielded a fascinating and well-preserved creature; Oreopithecus bambolii has skeletal features compatible with an upright posture and bipedal locomotion. Until the African Miocene yields something more appropriate, Oreopithecus is a candidate for a common ancestor, and interesting in another respect. Its dentition does not include prominent canine teeth that in the predominantly vegetarian, though occasionally carnivorous, Pan species serve well in their aggression-based, hierarchical social systems, as they do in the even more spectacular baboons.
Christopher Boehm, primate behaviouralist cum anthropologist, in his recent book Moral Origins (2012 Basic Books, ISBN-13: 978-0465020485) uses the principle of parsimony to reconstruct the social system of the Miocene Pan-Homo common ancestor from those of chimps and surviving human hunter-gatherers. His thesis is that it was centred on the hierarchical dominance of ‘alpha’ males, as is that of chimps. Prolonged social selection in hominin evolution largely tempered such a ‘Big Man’ tendency through a variety of strategies directed by majorities. Social punishments, including capital punishment, evolved to combat free-loading, theft and individual dominance in favour of cooperative egalitarianism. Such measures developed increasingly conscious self-suppression of such traits that eventually manifested themselves as what we now regard as human morals. Boehm considers that this psychological trend in evolution accelerated once Homo sapiens began hunting of large prey animals that added substantially to diet.
There is a major problem for this view: like Oreopithecus every well-preserved hominin species, even the earliest Sahelanthropus tchadensis, do not have prominent canines irrespective of whether they show evidence of at least partial meat-eating or pure vegetarianism. For some species with many fossil members, such as Au. afarensis, there are signs ofsexual dimorphism – larger males than females – but that does not necessarily signify hierarchical social behaviour. With the appearance of H. erectus that difference wanes to the present slight differences between modern male and female humans.
Agressive male humans, note gumshields (credit: John_Scone via Flickr)
If it is valid – and who knows? – for morphology to give clues to social behaviour, then it is equally likely that the beginnings of the hominin evolutionary thicket may well have involved a trend in social behaviour towards cooperative action; 8 million years ago. For generally small, gracile creatures with habits no more threatening to the big predators of the African savannahs that that of the porcupine, there would have been a powerful selection pressure towards a united front. Of course, in the last ten thousand years since the shift to economic strategies based on storable surpluses and their expropriation, hierarchical social systems with violence at their heart emerged among modern humans. Judging by the body shapes and dentition of extant ‘alphas’, as in capital’s boardrooms and among the frontbenchers at Westminster, anthropology clearly is in need of some refinement…
A means of assessing the cognitive abilities of hominins is through the objects that they created, whether tools or artefacts with apparent symbolic significance. The latter include pigments, coloured shells, beads, artwork or even deliberately parallel and crossing lines gouged on otherwise innocuous rock. Undoubtedly valuable to their creators, possibly treasured and passed on until lost or broken – most are fragile – symbolic artefacts are rare. So although they shout ‘thoughtful’, their age tells us little about when such a capacity first arose. Many archaeologists and palaeoanthropologists assert that creating and/or manipulating symbols may signify a link with being able to speak. Tools are a lot easier to find, probably as discards and lost items, and a well-described and understood sequence of forms and sometimes uses has been established, which extends as far back as perhaps 3 Ma – before the genus Homo appeared.
In terms of their meaning in terms of the consciousness of their makers and users, there are possibly four major recognisable steps. Chimpanzees and some birds can learn to pick up natural objects, such as stones and twigs, and use them: some bands of chimps even retain the knowledge. A step beyond that is preparing a natural object for use, as with breaking a pebble to create a cutting edge: something not exclusively human because it is possible that pre-human hominins created the earliest such Oldowan tools. Being able to visualise hidden potential inside something natural is altogether more advanced, and is represented by the iconic bi-face or Acheulean ‘hand-axe’. Its earliest makers, H. ergaster and erectus, literally brought such objects to light by skilfully knapping away the outer parts of substantial lumps of suitable rock. The knowledge endured for more than a million years but was eventually added to and superseded by a range of more delicate and specific stone tools, but more sophisticated tools represented the same ‘liberation’ of a simple idea held in rock. The fourth general cognitive leap was to add several resources together as composite tools, and arguably we have not long emerged from that phase with the creation of composite tools that help us design and make other tools: a machine-tool culture.
Example of a microlith (credit: Wikipedia)
It is that penultimate step-up in consciousness that has been engaging archaeologists since they first realised that some small, sharp chips of stone were not waste but deliberately crafted for combination with wood or bone. Such ‘microliths’ have been found in intact arrows and sickles of the Meso- and Neolithic, but their range steadily goes back in time with more research. Unmistakeable microliths have now been discovered at the South African coastal site at Pinnacle Point, in an occupation layer that is 71 ka old (Brown, K.S. and 8 others 2012. An early and enduring advanced technology originating 71, 000 years ago in South Africa. Nature, v. 491, p. 590-593).
The Pinnacle Point technology was indeed sophisticated, microlith manufacture requiring fire treatment as well as choice of rock and careful shaping and sharpening. As well as extending the microlith culture back so far the team of South African, US, Australian and Greek archaeologists compared them with 28 later African tool kits. The designs have barely changed from 71 ka to those of the last few hundred years. Kyle Brown and colleagues show that the industrial method endured, thereby laying to rest the somewhat reactionary notion that the methods were lost again and again in Africa after separate inventions and were only taken up decisively by the supposed ‘advanced’ anatomically modern humans who colonised Europe…
It is difficult to see how the Pinnacle Point microliths could have been useful, unless hafted in arrows or throwing sticks – maybe even saws and sickles? Crucially, they predate larger blade-tools that could have been hafted to form spears. The focus must now shift to the Zambian scene where possible microliths are reported at two 250 ka sites. If confirmed, they would link the decisive fourth cognitive step towards humanity with the very origin of fully modern humans, rather than a much later, non-African dawning of ‘smarts’ along with language, advanced art and much else in the chilly caves of southern Europe.
Of all human-colonised continents Africa lags far behind the rest as regards spread and density of archaeological digs. Only the ‘famous’ sites attract resources for investigation. Imagine what might emerge once there are more local people with research skills, equipment and transport; and, dare I say it, more independence of action and the attendant confidence in their ability.
Related articles
McBrearty, S. 2012. Sharpening the mind. Nature, v. 491, p. 531-532.
Male human head louse, Pediculus humanus capitis (credit: Wikipedia)
Until recently humans and lice were inseparable and the same goes for all primates, and nearly all mammals. However, unlike fleas, which happily will suck any blood that is going provided it is easily tapped, lice are tailored to their hosts. Should a baboon louse, for instance, get into your short and curlies it will almost certainly die. In any case, again unlike fleas, the louse cannot leap: they spread through intimate contact. The human head louse spreads especially well among nursery- and infant-school children, as any parent knows, because lessons often involve them literally getting their heads together. Less well known is that Pediculus humanuseschew soiled or greasy hair and it is the well-scrubbed kids who suffer and spread ‘beasts on the head’. Conversely, the clothes louse that carries typhus and other infections is deterred by regular laundry and ironing. And then there is the Continue reading “Hominin round-up”→
The Malapa valley South Africa, where Australopithecus sediba was found. (Credit: Lee R. Berger via Wikipedia)
The first stone tools and bones that had been cut by them, found in rocks dated at 2.5-2.6 Ma in the Bouri area of Ethiopia’s Afar Depression, have generally been taken as a sign that their invention was connected with more easily accessing meat for food. A corollary of this idea is that it was the introduction of meat into the hominin diet that helped ‘fuel’ the growth of their brains: meat-tools-brain interrelated in an evolutionary sense. There is a spatial link between such tools and fossils of Australopithecus, but direct attribution of the tools to these australopithecines has not been widely accepted. It is more generally accepted that a link to tools can be made with Homo habilis, but they lived, at the earliest, 200 to 300 ka later. The wear patterns on their teeth and association with animal bones bearing cut marks has been taken to indicate that at least part of their diet was meat.
Another approach to diet is to analyse the proportions of stable carbon isotopes (13C and 12C) in tooth enamel, which can discriminate between the ultimate plant source in their diet, i.e. between grasses that use the C4 photosynthetic pathway and the C3 version used by woody and herbaceous plants. The isotopic ‘signature’ of plants is also passed on to animals, depending on what vegetation they eat, and so up the food chain to predators and the scavengers that depend on their leavings. South African Au. africanus of around 2.5 Ma ago show a definite C4 preference as do local paranthropoids (‘robust’ australopithecine-like creatures) from around 1.8 Ma. The early humans H. habilis and H. ergaster also show the C4 isotopic proportions, which in both cases may be from a meaty diet or from a vegetarian component. The main point from these similar results, whatever the plant-meat proportions being consumed, is that these hominins were very different from chimpanzees in their eating habits, and probably as regards their habitats: savannah rather than woodlands respectively.
There are no reports of C-isotope research on Au. garhi teeth, but results from 2 Ma old Au. sediba found in South Africa have just been published (Henry, A.G and 8 others 2012. The diet of Australopithecus sediba, Nature, v. 487, p. 90-93) along with plant materials from dental plaque and tooth wear patterns. Au. sediba is notable for its very modern-looking hands and other ‘advanced’ features. Some believe it to have been closer to the direct line of human descent than a number of other hominin species, including the poor quality remains of H. habilis. So, did sediba eat meat? The forensic evidence suggests something unexpected. The C-isotope data points towards food dominated by C3 plants – less grasses and sedges, and more shrubbery. Tooth wear suggests bark was eaten, while plant remains from plaque indicate fruit leaves and wood. This is a feeding pattern more like that of chimpanzees than Homo species, Au. africanus and the paranthropoids that are roughly contemporary with Au. sediba. Ecological analysis of the sediments which buried the hominin specimens suggest a seasonal climate and savannah biome with abundant C4 plants that supported grazing herds, mixed with possibly some denser woodland along drainages. This is a pattern familiar from living savannah chimpanzee bands.
The hand and forearm of Australopithecus sediba (Credit: Peter Schmid, courtesy Lee R. Berger via Wikipedia)
So, despite being an ‘advanced’ hominin, by carrying clear signs of foods that were not consumed by meaty potential prey animals Au. sediba probably was not a meat eater. Yet species with strong C4 ‘signatures’ cannot be assigned to carnivory on C-isotope evidence alone. One has to decide from other data, such as tooth-wear and plaque, whether this or that hominin ate grasses, those that clearly did not becoming candidates for dominantly meat-eating. How to detect a tool-using species with a mixed diet, akin to more modern humans, is a tough nut to crack.







 left, Lucy (AL...")



")













